1Yunnan Provincial Key Laboratory of Public Health and Biosafety, School of Public Health, Kunming Medical University, Kunming 650500, China.
2The Institute for Tropical Medicine, School of Basic Medical Sciences, Kunming Medical University, Kunming 650500, China.
3Yunnan Province Key Laboratory of Children’s Major Diseases Research, The Affiliated Children’s Hospital, Kunming, Kunming Medical University, Kunming 650030, China.
4Yunnan Demonstration Base of International Science and Technology Cooperation for Tropical Diseases, Kunming 650500, China.
Aihua Liu & Fukai Bao
Email: liuaihua@kmmu.edu.cn & baofukai@kmmu.edu.cn
Received : Nov 29, 2023 Accepted : Dec 18, 2023 Published : Dec 28, 2023 Archived : www.meddiscoveries.org
Borrelia burgdorferi Basic membrane protein A (BmpA) plays a crucial role in mediating immune responses to Lyme arthritis. The aim of this study was to describe manifestations and the main evaluation method on recombinant BmpA (rBmpA) - induced arthritis in tree shrews. We used rBmpA to stimulate knee joint in vivo in tree shrew animals. Arthritis was evaluated by Arthritis index, pathology, expression of proinflammation cytokines. Results showed that the joints of tree shrews appeared swelling, both the scores of arthritis and the number of swelling joints were enhanced The levels of TNF-α and CXCR4 increased in Peripheral blood. Day 7-15 after BmpA injection, the inflammation reached its peak. Histopathology examination showed that the synovium was hyperplasia, arranged irregularly. Our data indicated that the rBmpA-induced tree shrew arthritis may be used as a candidate model for Lyme arthritis pathogenesis and treatment.
Keywords: Lyme arthritis; Borrelia burgdorferi; rBmpA; Tree threw.
Lyme Disease (LD), a serious inflammatory syndrome caused by the spirochete Borrelia burgdorferi (B. burgdorferi, Bb), is characterized by tissue inflammation of the skin, muscles, joints, and central nervous system (CNS). Lyme disease occurs throughout North America, Europe (particularly central Europe and Scandinavia, Russia), and Asia [1]. Since Lyme disease was recognized in 1976 [2], 300,000 cases have been reported annually in the United States [3,4].
Lyme arthritis is inflammatory arthritis that is likely mediated by the immune response to the spirochete in the joint and can be seen in the early or late stage of Lyme disease [5]. Lyme arthritis has been characterized as a mono- or oligoarthritis, typically presenting as a monoarthritis of the knee [6,7]. Patients with Lyme disease could suffer from varying degrees of pain and swelling of the knee as well as skin involvement with redness and oedema around the ankle of the same leg [8,9].
BmpA is a major membrane protein that is closely associated with Lyme arthritis [10]. BmpA, a specific, highly conserved, and immunogenic lipoprotein, is encoded by the bmp family genes of paralogous chromosomal gene family 36 [11,12]. BmpA operons that are induced in murine and human joints possess inflammatory properties. BmpA-deficient B. burgdorferi persists poorly in joints and fails to induce arthritis compared to the wild type B. burgdorferi [13]. An isogenic BmpA mutant induced significantly lower levels of pro-inflammatory cytokines TNF-alpha and IL-1beta in cultured human synovial cells. BmpA can induce cytokine responses in synovial cells via activation of the NF-κB and p38 MAP kinase pathways [14].
In the past 45 years, a wide variety of animals have been used to study the pathogenesis of Lyme arthritis and determine therapeutic targets. These animals include rodents [15] that are most used (C3H/HeN mice, C57BL/6J mice, Gamma interferon gene-deficient mice, C3H/HeN-SCID mice, BALB/C mice), hamsters and nonhuman primates [13,16-22]. Although the current infection models of Lyme arthritis are useful in understanding the basic immunological events that lead to the development of Borrelia burgdorferi-induced inflammation, these findings need confirmation in humans [23]. High costs limit the routine use of large non-human primates. The Northern Tree shrew (Tupaia belangeri) is a small squirrel-like mammal indigenous to southwest Asia. Compared with rodents, the Tree shrew genome shows higher similarity to humans, and many of their biological characteristics have close homology with humans [24]. Importantly, from the perspective of infectious diseases, tree shrews are susceptible to infection with human pathogens and manifest clinical signs akin to human infections [25,26]. Therefore, using Borrelia burgdorferi bmpA to induce the occurrence of arthritis in tree threw could be a better choice.
Ethics statement
Tree shrews were obtained from the Kunming Medical University, Yunnan, China*
(https://www.kmmc.cn/list1370.aspx).
The animals were bred and maintained at the Animal Laboratory Center of Kunming Medical University in strict accordance
with the rules and regulations of China animal welfare law.
All animal experiments were approved by the Experimental
Animal Committee of Yunnan (Permit Numbers: LA2021744, JXJY20200018).
rBmpA preparation
E. coli expression system was used to produce recombinant BmpA (rBmpA) using the bacterial expression plasmid vector pGEX- 6P-1 (GE, USA) (Figure 1), and the following BmpA primers was used (5′-3′containing EcoRI and Xhol restriction sites): forward, ACG AAT TCA TGA ATA AAA TAT TGT TGT TGA, reverse, AGC TCG TAA ATA AAT TCT TTA AGA AA. Expression, purification, and enzymatic cleavage of the glutathione transferase fusion protein were carried out as described before [10].
Animal preparation
Thirty-two adult female Tree shrews (4-6 months old, 150 g mean weight). Animals were routinely fed in a isolated cages in an experimental animal room under the following conditions: 25±2°C temperature, 50% relative humidity, natural ventilation 8~10 h/d, noise <60 dB, and 12h/12h light/dark cycle.
Induction of arthritis
The purified rBmpA protein was diluted with 0.01M PBS and injected into the left knee joints of tree shrews following different concentrations: Low dose groups (0.04 mg/ml, 50 µL), high dose groups (0.08 mg / ml, 50 µL), PBS groups (0.01M, 50 µL). All tree shrews were then examined daily for joint swelling and assigned a clinical arthritis score (Figure 1).
Arthritis scoring
The arthritis scores were scored twice a week by two people who were not sure about the experimental group. Arthritis index score criteria (0-4) were as follows [27]: 0, no swelling or inflammation; 1, slight swelling and/or redness of paw; 2, moderate swelling in joints; 3, pronounced swelling with more than two joints; and 4, severe swelling with joint rigidity. The arthritis score was the sum of the affected joints in each tree shrew’s hind limbs.
Tissue collection and histopathologic analysis
The tree shrews in each group (n=8) were euthanized by CO2 asphyxia on the 28th day after injection. The hind limbs of each animal were immediately fixed in 10% formalin, then decalcified, implanted with paraffin, sectioned and stained with haematoxylin-eosin. The slides were observed under high power microscope (×40) taken photos.
Tree shrew blood cytokines detection
The heart blood was collected from Tree shrews on the 7th, 14th, and 21st days after the injection. Total RNA was extracted from whole blood using RNAiso Plus reagent (TaKaRa). A Prime Script RT reagent Kit with Gdna Eraser (TaKaRa) was used to synthesize cDNA, according to the manufacturer’s protocol. Realtime PCR was conducted using SYBR Premix Ex Taq (TaKaRa), according to the manufacturer’s protocol. Relative gene expression levels were calculated using the comparative threshold cycle method for quantitative PCR, with data normalized to the housekeeping gene GAPDH. Sequences of the specific primers used to amplify GAPDH, TNF-α, IL-2, IL-6, IL-10, IL-17, CXCR4 are listed in Table 1.
| Gene | Forward primer (5´→3´) | Reverse primer (5´→3´) |
|---|---|---|
| GAPDH | GGTGCCGAGTATGTTGTGGA | TCTCATGGTTCACGCCCATC |
| TNF-α | CGCTACCTGGACTTTGCTGA | CCCTGAGCCCCTAATTCCCT |
| IL-2 | CGTCACAAACAGTGCACCTAC | CCTGGAGAGCATCTTTGGGTT |
| IL-6 | CGTCACAAACAGTGCACCTAC | CCTGGAGAGCATCTTTGGGTT |
| IL-10 | AAACCAAACCACAAGGGCAC | AGGTTGGCTGGGAAGTAGGT |
| IL-17A | CTGTGTGGATGCTGAAGGGA | CAGCCCACAGTCACCAGTAT |
| CXCR4 | GACTTGTGGGTGGTAGTGTTCC | GACTGTGGTCTTCAGGGCTTTG |
Statistical analysis
All data are shown as means ± Standard Deviation (SD). Statistical analysis were performed using GraphPad Prism 9.2.0 software (GraphPad Software Inc, San Diego, CA). Two-way Analysis Of Variance (ANOVA) was used for comparison of multiple groups, followed by Tukey’s multiple comparison test for post hoc test. P<0.05 was set as the level of statistical significance.
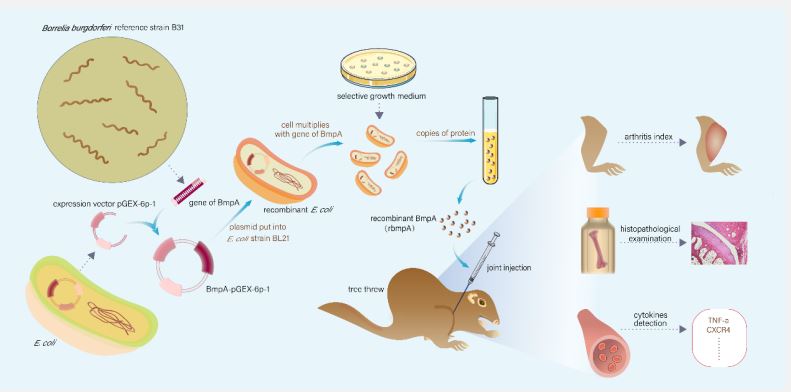
Animal observation
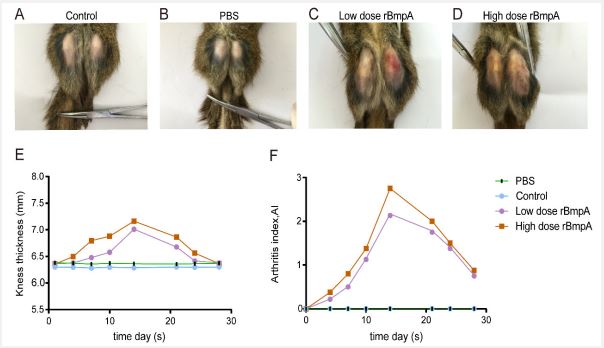
The tree shrews in the healthy control group (Figure 2A) and the PBS control group had normal feeding, flexible activities, no “wet tail” caused by diarrhea, no bleeding at the injection site, no rash, and slight redness and swelling at the local injection site 24h after the injection lasted only 2 to 3 days. No other abnormality post-injection was seen by the naked eye (Figure 2A,B). In experimental group, food intake, activity, and body response of the tree shrews were generally reduced on day 7 after rBmpA injection, which manifested as often animals hiding in cages and not easy to be driven.
The degree of joint swelling in rBmpA-injected tree shrew group
The knee joint swelling and congestion in the low-dose rBmpA group were all obvious. The knee joint swelling in the highdose rBmpA group was more clear, but the congestion was not obvious (Figure 2C,2D). Both rBmpA groups reached the peak of lesions in 7-15 days, and then the symptoms gradually alleviated after 3 week (Figure 2E). rBmpA groups have small ulcers on their knee joints after rBmpA injection, and the scabs can be repaired in about 2-3 days. The stool under the cage was observed to be green and loose. After anatomy, it was found that the intestines were dark. It may be caused by enteritis, diarrhea, and dehydration. Our evidence showed that there were no obvious lesions on the front paws, tails, and other surfaces of the tree shrews in each group. One week after the injection, the incidence of arthritis in the experimental group reached 80%, and the arthritis score was 2-6. The average arthritis index reached a peak two weeks after the injection (Figure 2F). The clinical symptoms in the high-dose group are easier to observe. The tree shrews in the health control group (Figure 2A) and PBS group (Figure 2B) showed no swelling and congestion of hind limb.
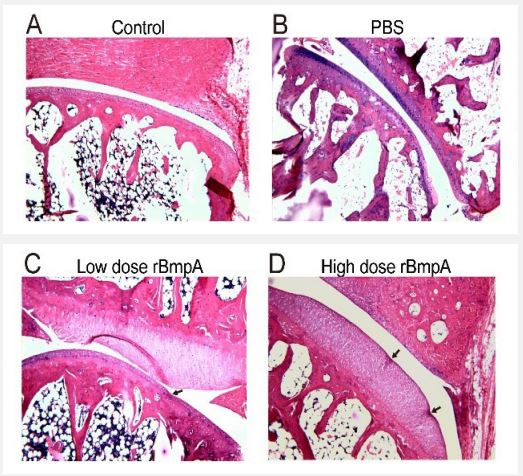
Injection of rBmpA triggered synovial cartilage degeneration
All tree shrews were euthanized on day 28 postinjection. Both hind limbs of each euthanized animal were immersionfixed, decalcified, and stained with hematoxylin-eosin for histopathological evaluation. Tree shrews in the rBmpA groups (Figure 3C,3D) showed early manifestations of cartilage degeneration, with normal joint spaces, irregular articular surfaces, depressions in the synovial cartilage of the joints, and irregular cell arrangement in the depressions. In addition, the irregular depressions or line-like grooves on the surface of articular cartilage showed chondrocyte proliferation and cartilage microfibrosis. Such articular cartilage degeneration were absent in control group injected with PBS (Figure 3B). Our data indicate that local exposure of the joints to rBmpA is sufficient to trigger the early pathological changes of arthritis.
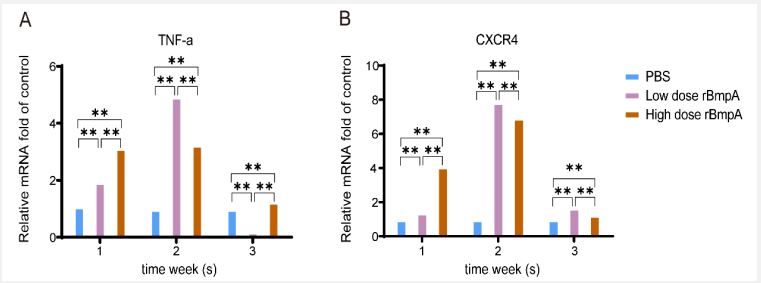
The expression of TNF-α and CXCR4 in tree shrews
To further study pathogenesis of rBmpA-induced arthritis, the expression levels of IL-2, IL-6, IL-10, IL-17A, TNF-α, and CXCR4 mRNA in peripheral blood of tree shrews were determined by RT-PCR following rBmpA injection. Compared with the untreated control group, TNF-α and CXCR4 mRNA expression levels in tree shrew peripheral blood increased significantly after rBmpA stimulation for 1,2, and 3w, reaching a peak at 2 weeks. However, there were no significant differences in IL-2, IL-6, IL-10, and IL-17A mRNA expression levels in experimental group relative to the control group at any time point.
The pathogenesis of Lyme arthritis is still under exploration. Our study confirmed that using rBmpA to inject into knee joint of tree shrews can induce Lyme arthritis.
Tree shrew as an animal model broadens the limitations of Lyme arthritis research. Since tree shrews are closer to humans, we used tree shrews to simulate the characteristics of human Lyme arthritis. So far, Borrelia burgdorferi has been used to infect variety of animal models, including rodents [28-30], rabbits [31,32], dogs [33,34], monkeys [35] horses [36], cattle [37] and people, etc [38]. However, the animal models for Lyme arthritis are mainly mice. These mice provide important insights and valuable knowledge for understanding the pathogenesis of Lyme arthritis. Although mice are widely used because of their small size and clear genetic background, there are significant genetic differences between rodents and humans. Be cautious when translating these findings directly into the human system [39]. Mice sets limitations for the study of human Lyme arthritis. Non-human primates are genetically close to humans, and they can simulate human physiology and pathology relatively accurately. However, their high cost and low productivity also severely limit their use. Human body experiments, due to issues related to the maintenance of human ethics, and the difficulty of recruiting volunteers, hinder the possibility of its implementation. Compared with other experimental animals, tree shrew genome sequencing showed that its neural, immune, and metabolic systems are very close to humans, and it is a small mammal closely related to primates. It has been proposed as an alternative laboratory animal to primates in biomedical research [40].
Our research is to use rBmpA instead of live Borrelia burgdorferi as an inducer for its close relationship with Lyme disease (Figure 1). At present, the establishment of animal models of Lyme arthritis is mainly carried out by subcutaneous injection of live Borrelia burgdorferi [41]. This induction method is intended to simulate the natural transmission method of tick bites to infect animals, but some animal studies also used needle inoculation instead of tick transmission to infect animals, compared to the natural method of tick bites. It have found that, For spirochetes that enter animals in this way, the eradication of Borrelia burgdorferi with antibiotics may not be complete. Borrelia burgdorferi DNA can be detected in animals tissues up to 9 months after antibiotic treatment [34,35,42-44]. In the past, it was believed that if this kind of tissue residual spirochetes exists in humans, it is the cause of post-Lyme disease syndrome (persistent musculoskeletal pain, neurocognitive symptoms, or fatigue feelings after antibiotic treatment, etc.) [45-47]. However, the current view believes that there is a serious methodological problem that does not consider the pharmacokinetics of antibiotics when choosing the dosage regimen used Kineticpharmacodynamic properties [48], the pathogenesis of later features of LD may no longer be dependent on infection [49]. The study found that BmpA is one of the exposed and highly antigenic proteins on the surface of Borrelia burgdorferi. BmpA can induce cytokine response in synovial cells. NF-κB and p38 MAP kinase pathway activate the pro-inflammatory response of human synovial cells [14]. In mice, it may trigger a complex cascade of host inflammatory responses that cause arthritis. It was found that the expression levels of Borrelia burgdorferi bmpA and bmpB were significantly higher than that of the skin, suggesting that bmpA may also be applicable to Lyme arthritis model [13]. Our previous studies have also shown that the basic membrane protein A of Borrelia burgdorferi triggers a storm of pro-inflammatory chemokines in THP1-derived macrophages through receptors TLR1 and TLR2, which may trigger innate and adaptation in infected arthritis [10]. Therefore, on the one hand, in order to avoid the controversy of Lyme disease syndrome caused by residual spirochetes, we can use rBmpA to replicate Lyme arthritis model [13,14,50].
rBmpA induces pathological changes in the synovium of tree shrew joints (Figure 3). The pain caused by human Lyme arthritis can be seen in the early or late stage, the early stage is mainly proliferative synovitis [51], and some patients in the late stage can develop chronic and severely destructive Lyme arthritis [8], and its pathological characteristics include synovial hyperplasia, infiltration of monocytes into the synovium, formation of inflammatory exudates on the synovial cell wall (vascular loop formation), cartilage destruction, and bones and joints [52,53]. C3H mice, the most commonly used animal models for Lyme arthritis [23], whose severity of this arthritis usually peaks around 3 weeks after the Intradermal inoculation of susceptible strains, after which the pathological changes quickly subside. In addition, joint edema changes are usually accompanied by histological changes. In the infection model of Lyme arthritis mice, this swelling can be observed about 2 weeks after infection and gradually decreases. In our study, the degree of joint swelling and arthritis index of tree shrews reached their peak at 2 weeks after being infected with rBmpA, and then gradually decreased (Figure 2). The pathology of the tree shrew’s joints is that the early manifestations of cartilage degeneration appear after 4 weeks (Figures 3C and 3D).the articular surface is irregular, the joint synovial cartilage is recessed, the cell arrangement of the recessed area is irregular. Because it is a preliminary exploration, the observation time is also being explored. If continuous long-term observation is continued, further research can be done on whether rBmpA can cause persistent Lyme disease arthritis.
In this study, we found that increased TNF-α (Figure 4A) and CXCR4 (Figure 4B) in the blood of the tree threws in response to rBmpA of Borrelia Spirochetes. An previous study reported that rBmpA initiates proinflammatory chemokine storm in THP 1-derived macrophages in humans via the receptors TLR1 and TLR2 [10]. TLRs are a conservative molecular model in which the receptors of the host cell can recognize microbial components such as lipoproteins, lipopolysaccharides, flagellin, and nucleic acids [54]. The rBmpA used in our study is one of the lipoprotein molecules on the surface of the Borrelia burgdorferi membrane, which may be recognized by TLRs to initiate the innate immune response. Studies have shown an increase in transcript levels of genes encoding CC and CXC chemokines, proinflammatory cytokines, and TNF superfamily members in the joint tissues infected with live Borrelia burgdorferi-infected mice [55-57]. Under the stimulation of Borrelia burgdorferi, neutrophils promote the development of arthritis by releasing pro-inflammatory cytokines, including TNF-A, IL-1, IL-8, IL-15, and so on [58,59]. Chemokines can regulate leukocyte trafficking during host defense immune response [60]. CXCR4 is one of the receptors of the chemokine subfamily CXC, a typical G Protein-Coupled Receptor (GPCR) [64,65], which can activate Gprotein-mediated downstream signaling pathways [66], whose ligand is CXCL12, an important member of the CXC chemokines [16]. The CXCL12-CXCR4 axis is well established to play multiple roles in cellular migration, survival, and proliferation, as well as in other functions [61-63]. CXCL12 and CXCR4 were highly expressed in the serum and joint synovial fluid of patients with Rheumatoid Arthritis (RA) [64]. CXCR4 levels in synovium have been correlated with joint destruction in RA patients [65]. The subset expressing CXCR4 in the peripheral blood positively correlated with TNF-α in RA patients [66]. TNF-α has involved the occurrence of inflammation and the pathogenesis in RA [67,68]. TNF-a and CXCR4 reached their peaks 2 weeks after rBmpA inoculation (Figure 4), which was consistent with the process of joint swelling (Figure 2). Our experiment suggests that CXCR4 and TNF-α may be involved in the inflammation of Lyme arthritis.
Moreover, studies have shown that CXCR4 was both relatively highly expressed in the spleen and thymus of tree threws, which implying that CXCR4 may be involved in some aspect of immunological regulation among tree shrews. The study showed that CXCL12 and CXCR4 in tree threws are structurally similar to their homologous human proteins. The similarities between tree shrew and humans CXCR4 were as high as 97%, and those of mice and rats were 91% and 92%, respectively [69]. This shows that the expression of CXCR4 in tree shrews is closer to humans than in mice and rats. Tree shrews might be more suitable for explaining human clinical phenomena. Our experiment implicated that tree shrews CXCR4 level increase during joint swelling (Figure 2) may play similar roles as those in humans Lyme arthritis. Tree shrew may be a viable animal model for studying human Lyme arthritis.
In the next step, we plan to extend the observation time of tree shrew model, increase the induction mode of rBmpA and use more experimental verification methods which is beneficial to the research of Lyme arthritis.
Our study identifies rBmpA as a likely contributor to inflammatory responses in vivo in tree threw arthritis. Persistence of this antigen in the joint may lead to synovitis. Using new method of to simulate Lyme arthritis of tree shrews with rBmpA immunogen helps to expand Lyme arthritis model spectrum. The expression of tumor necrosis factor-α and CXCR4 gene in whole blood of tree shrews increased significantly and could correlate to Lyme arthritis pathogenesis. Manifestations in tree shrew arthritis suggests a potential role of rBmpA in the pathogenesis of Lyme arthritis.
Acknowledgements: This work was supported by grants from the National Natural Science Foundation of China ( No. 32060180, 82160304, 81860644, 81560596 ), Joint Special Project of Yunnan Science and Technology Department and Kunming Medical University [No.2019FE001 (-002) and 2017FE467 (-001)]. The funding institutions had no involvement in the design of the study or review of the manuscript.
Author contributions: Fukai Bao, Aihua Liu, and Bingxue Li conceived and designed the experiments. Yuxin Fan, Shiyuan Wen developed the methodology. Bingxue Li, Hanxin Wu, Weijie Ma, Lei Zhong, Bingxue Li, Li Peng, Li Gao, Xinya Wu, and Weijiang Ma performed all experiments. Weijiang Ma, Liangyu Zhu, Jieqin Song, Suyi Luo analyzed and discussed the data. Bingxue Li wrote the manuscript. Fukai Bao, Aihua Liu, and Bingxue Li edited and revised the manuscript. All authors read and approved the final manuscript.
Competing interests: The authors declare no competing interests.
