1College of Veterinary Medicine, Northeast Agricultural University, Harbin, 150030, China.
2School of Life Sciences, Northeast Agricultural University, Harbin, 150030, China.
3College of Food, Northeast Agricultural University, Harbin, 150030, China.
4College of Engineering, Northeast Agricultural University, Harbin, 150030, China.
Shichao Liu
Tel: 18545078137;
Email: LSC10251025@163.com
Received : Oct 28, 2024 Accepted : Nov 18, 2024 Published : Nov 25, 2024 Archived : www.meddiscoveries.org
Emerging evidence reveals an important role of Heat shock protein 90 kDAα, Class B, Member 1 (HSP90AB1) in cancer initiation, development, and progression, but no pan-cancer analysis of HSP90AB1 has been performed. In this study, the expression profile and prognostic significance of HSP90AB1, as well as the relationship between HSP90AB1 and clinicopathological parameters and immune cell infiltration were comprehensively studied by bioinformatics technology. HSP90AB1 is significantly up regulated in a variety of common cancers and is associated with prognosis. Cox regression analysis showed that HSP90AB1 had high expression of bladder unessential carcinoma, cholangiocarcinoma, lung adenocarcinoma in Overall Survival (OS), Disease-Specific Survival (DSS), and Progression-Free Interval (PFI) were associated. In most cancers, HSP90AB1 DNA methylation is reduced and inversely correlated with HSP90AB1 expression. Increased phosphorylation in HSP90AB1 has been complied with in some cancers. HSP90AB1 was substantially correlated with infiltrating cell and immune checkpoint gene levels. Our pan-cancer analysis provides a multifaceted understanding of the function of HSP90AB1 in tumorigenesis and metastasis in different cancers.
Keywords: HSP90AB1; Pan-cancer analysis; Prognostic biomarker; Immune cell infiltration; Cancer immunotherapy.
Cancer, the major cause of morbidity and mortality worldwide, imposes major healthcare-linked economic burdens on society [1]. Countless oncogenes and tumor suppressor genes have been determined, which are related to signaling pathways that dominate the growth or death of cell. This enabled the exploration of anti-tumor agents and therapeutic techniques. However, there is still a long way to move to eradicate cancer due to the increasing drug resistance [2]. Therefore, there is urgent need to identify new diagnostic biomarkers and therapeutic targets [3,4].
The Heat Shock Protein (HSP) family consists of 95 revolutionising conserved genes, which play a key role in protein homeostasis during normal and cancerous processes [5]. Heat Shock Proteins (HSPS) are a group of proteins produced by cells under the induction of stressors, especially environmental high temperature, and are highly conserved in the evolutionary process. It can improve the stress ability of cells, especially the heat resistance, and also enhance the resistance to various injuries. Studies have shown that the mass production of heat shock proteins in cancer cells can promote the growth of cancer cells, and is closely associated with the occurrence, development and deterioration of cancer [6].
In recent years, HSP90AB1, as a novel biomarker, has been widely used in drug screening and biomedical research. HSP90AB1 has high expression and stability, and can be used as a protein biomarker for disease diagnosis and treatment. In the aspect of drug screening, HSP90AB1 has excellent selectivity [7]. Numerous drugs, such as anti-tumor drugs, anti-infective drugs, cardiovascular drugs, etc., have different targets. By binding to these drugs, HSP90AB1 can generate specific fluorescence signals, providing an important basis for drug screening. Based on RNA sequencing data, HSP90AB1 was identified as the most promising target among the other predicted HSPS [8]. In addition, HSP90AB1, which is highly expressed in a variety of cancers, also promotes tumor formation and cancer cell proliferation, migration, and glycolysis, and goes hand in hand with poor prognosis. As such, they are also targets for innovative treatments for cancer treatment [9].
At present, there are few studies on the occurrence and progress of HSP90AB1 in cancer, and most of them are aimed at a specific cancer. We performed the pan-cancer analysis of prognosis and immunity HSP90AB1 in detail and systematically with the help of a database to determine its feasibility as a innovative cancer therapeutic target.
With the development of bioinformatics and public databases, pan-cancer analysis and its relationship with clinical prognosis have become a topic of interest to researchers, and have become an significant means to elucidate the possible molecular mechanisms of oncogenesis [10]. In this study, we conducted a comprehensive systematic review of the role of HSP90AB1 in numerous cancer types, including its expression, function, and prognostic value. We also investigated the association of HSP90AB1 with tumor microenvironment, immune cell infiltration, and other immune-related biomarkers based on an online web server and R program. The study highlights the potential and critical role of HSP90AB1 as a biomarker for prognosis and immunotherapy in pan-cancer, setting the stage for subsequent prospective studies as well as function and mechanism.
Data collection and differential gene expression analysis
Clinical follow-up information for 33 cancer Types of the Cancer Genome Atlas (TCGA) from UCSC database (https://xenabrowser.net/datapages/) was download. The sequencing data of HSP90AB1 also came from the GTEx project. The red and blue boxes represent tumor and normal tissue, respectively. The expressed data was transformed by log2 (TPM+1) to eliminate missing data and duplicate values. ∗p<0.05, ∗∗p<0.01, and ∗∗∗p<0.001 (Figure 1A). The HSP90AB1 protein expression level in normal tissues and primary tissues of 8 of them were examined using the CPTAC dataset. We examined their Z-value with the help of CPTAC prodromic data, as well as the statistical significance between normal and primary.
Pathological stage analysis
Based on TCGA data, the correlations of HSP90AB1 expression with 19 major pathological stages, including ACC, BLCA, BRCA, CESC, CHOL, COAD, DLBC, ESCA, GBM, HNSC, KICH, KIRC, KIRP, LAML, LGG, LIHC, LUAD, LUSC and MESO were investigated. Log2 (TPM+1) was used for logarithmic scale. ∗p<0.05, ∗∗p<0.01, and ∗∗∗p<0.001.
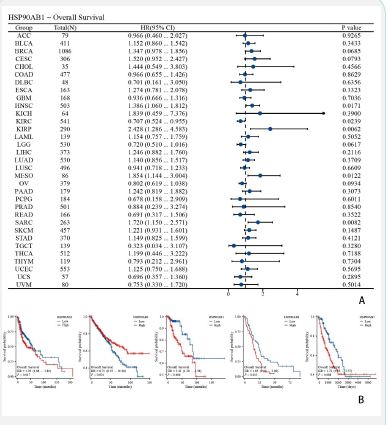
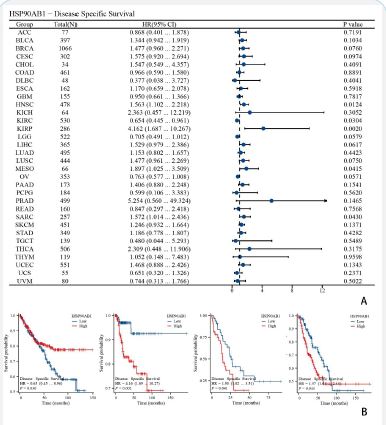
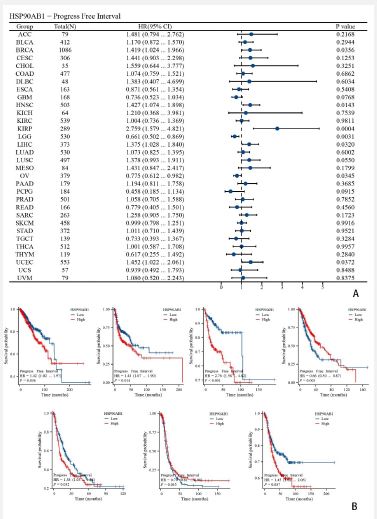
Analysis of the influence of HSP90AB1 expression on tumor OS and DSS
Forest plots showed the univalent Cox regression results of HSP90AB1 for OS, DSS and PFI in TCGA pan-cancer types. The effect of HSP90AB1 mutation status on overall survival of some kinds of cancer patients was investigated using the cBioPortal database (Figure 3A). Showed KIRP, LGG, LIHC, MESO and UCEC (Figure 4A) showed KIRP, LGG, LIHC and MESO (Figure 5A). Showed BRCA, HNSC, KIRP, LGG, LIHC, OV and UCEC.
DNA methylation and mutation characteristics analysis
UALCAN database and cBioPortal database were used to investigate DNA methylation and mutation characteristics of HSP90AB1. The promoter methylation levels of HSP90AB1 in primary tumor, including BLCA, LUSC, PAAD, COAD, PRAD, CESC, ESCA, KIRP, LIHC, BRCA, SARC, TGCT, UCEC and LUAD, were showed in (Figure 6A). Alteration frequency of mutation, amplification and deep deletion of 63 cancers were showed in (Figure 6B, Figure 6C). Compared the survival curves of ACC and BLCA patients both with and without genetic alterations in HSP90AB1.
Phosphorylation analysis of hsp90ab1 in cancer
The phosphorylation of HSP90AB1 in normal tissues and primary tissues of Ovarian cancer, Colon cancer, Clear cell RCC, Lung adenocarcinoma, Lung adenocarcinoma, Head and neck squamous carcinoma, Pancreatic adenocarcinoma, Glioblastoma multiflora and Hepatocellular carcinoma was examined using the CPTAC dataset.
KEGG and GSEA enrichment analysis of HSP90AB1
KEGG analysis was used to detect the biological and molecular functions of HSP90AB1. We also used GSEA to identify the potential molecular mechanisms of HSP90AB1. GSEA analysis showed the enrichment of HSP90AB1 in BP, CC, MF and KEGG respectively.
Pancarcinoma analysis of HSP90AB1 expression and immune cell infiltrations
The horizontal axis in the heat map represented different types of cancer, the vertical axis represented different immune scores, and the different colors represented correlation coefficients. Red meant positive correlation, and blue meant negative correlation ∗p<0.05.
Correlation analysis of HSP90AB1 expression and immune checkpoint gene in pericardium
Spearman heat maps of immune checkpoint related genes and HSP90AB1 gene expression in various cancers were generated. The horizontal axis in the heat map represents different types of cancer, the vertical axis represents different immune scores, and the different colors represent correlation coefficients.
Expression of HSP90AB1 in pancarcinoma
Kaplan–Meier curves showing the association between HSP90AB1 expression and Overall Survival (OS) for KIRP, LGG, LIHC, MESO and UCEC.

We used the TCGA database to analyze the differential expression of HSM1 between normal and cancerous tissues, including Adrenal Cortical Carcinoma (ACC), Bladder Unessential Carcinoma (BLCA), Breast Invasive Carcinoma (BRCA), cervical squamous cell carcinoma and cervical duct adenocarcinoma (CESC), Cholangiocarcinoma (CHOL), Colorectal Adenocarcinoma (COAD), symphonious Diffuse Large B-Cell Lymphoma (DLBC), and food Tubular carcinoma (ESCA), Glioblastoma Multifaceted (GBM), Head and Neck Squamous Cell Carcinoma (HNSC), chromophobe cell carcinoma of kidney (KICH), clear cell carcinoma of kidney and kidney (KIRC), papillary cell carcinoma of kidney and kidney (KIRP), Acute Myeloid Leukemia (LAML), Low Grade Glioma Of Brain (LGG), Hepatocellular Carcinoma (LIHC), Lung Adenocarcinoma (LUAD), Lung Squamous Cell Carcinoma (LUSC), Mesothelioma (MESO), ovarian serous cystadenocarcinoma (OV), Pancreatic Adenocarcinoma (PAAD), pheochromocytoma and paraganglioma (PCPG), prostate adenocarcinoma (PRAD), rectal adenocarcinoma (READ), Sarcoma (SARC), cutaneous melanoma (SKCM), gastric adenocarcinoma (STAD), Gastric and testicular germ cell tumors (TGCT), thyroid cancer (THCA), thymoma (THYM), Body endometrial cancer (UCEC), Uterine Sarcoma (UCS) and Uveal Melanoma (UVM). The results showed that HSM1 was substantially overexpressed in 16 cancer types. Specifically, in cancer types such as BLCA, BRCA, CHOL, COAD, ESCA, HNSC, KIRC, LIHC, LUAD, LUSC, PAAD, PCPG, PTAD, READ, SARC, SKCM, STAD, and THYM, its expression level in cancer tissues was significantly higher than that in neighboring non-malignant tissues, while the opposite pattern was found in UCEC (Figures 1A). Under the help of CPTAC proteome data, we focused on 8 of the 18 cancers and studied their Z-value (Figures 1B). We also compared the statistical significance between the normal and the primary. A table was drawn according to the results analyzed, through which we found that among the eight cancers listed, breast cancer showed the strongest statistical significance with HSP90AB1 expression, while the statistical significance of colon cancer was the worst.
Correlation between HSP90AB1 and pathological stage
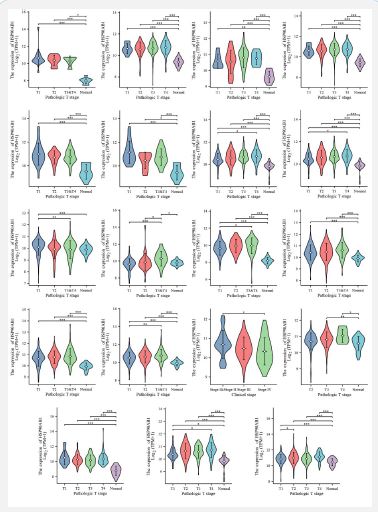
As is shown in (Figure 2). By evaluating HSP90AB1 expression in patients with stage I, II, III, and IV cancers, the association between HSP90AB1 expression and clinicopathological features in a variety of cancers can be studied. We evaluated in patients with stage I, II, III, IV cancers and normal people. The results of TCGA database showed that the expression of HSP90AB1 in most cancers, such as ACC, BLCA, CHOL, and COAD, was significantly different from the normal (Figure 2), and almost in all types of cancers, HSP90AB1 expression was higher in stage T1-T4 than normal. HSP90AB1 expression is consistent in cancer from stage T1 to stage T4, like BLCA, ESCA, LIHC, and LUSC. What’s more, HSP90AB1 expression showed no remarkable differences among several stages in most of the cancers studied.
Effect of HSP90AB1 expression on tumor OS and DSS
As is shown in (Figures 3-5). We performed survival association analyses (including OS, DSS and PFI) to determine the association between HSP90AB1 expression levels and prognosis for 33 cancer types. The results of OS analysis showed that HSP90AB1 was a risk factor in the patients with BRCA, CESC, HNSC, KIRP, MESO and SARC, and a protective factor in the patients with KIRC, LGG and OV (Figure 3A). As for DSS, the results of Cox regression analysis showed that HSP90AB1 was significantly correlated with worse DSS in HNSC, KIRP, LGG, LIHC, MESO and SARC, especially in KIRP, while it was associated with better DSS in KIRC and OV (Figure 4A). For PFI, worse PFI was linked to HSP90AB1 expression in BRCA, HNSC, KIRP, LIHC and UCEC, and better PFI was linked to HSP90AB1 expression in LGG and OV patients (Figure 5A).
K-M plotter analysis indicated that higher HSP90AB1 expression levels were connected with poorer OS in patients with KIRP, LGG, and UCEC, while lower expression levels were associated with worse OS in patients with LIHC and MESO (Figure 3B). On the other hand, K-M plotter analysis showed that elevated HSP90AB1 expression was related to worse DSS in patients with LGG, LIHC and MESO, while KIRP patients with higher HSP90AB1 expression had a longer DSS time (Figure 4B). What’s more, it’s found that higher HSP90AB1 expression levels may lead to lower survival probability and shorter survival period in KIRP, LIHC and UCEC, and lower HSP90AB1 expression levels were linked to lower survival probability or shorter survival period or both in BRCA, HNSC, LGG and OV (Figure 5B).
DNA methylation of hsp90ab1 in pancarcinoma and its effects
As is shown in (Figure 6), DNA methylation constitutes one of the forms of epigenetic inheritance that affects the occurrence and development of cancer directly. By using UALCAN database and cBioPortal database, we investigated DNA methylation and mutation characteristics of HSP90AB1 in pancreatectomy. Analysis of the data demonstrated that the promoter methylation levels of HSP90AB1 in primary tumor were higher than normal in BLCA, LUSC, PAAD, COAD, PRAD, CESC, ESCA, KIRP and LIHC patients. In contrast, lower HSP90AB1 promoter methylation levels were discovered in patients with BRCA, SARC, TGCT, UCEC and LUAD (Figure 6A). By utilizing cBioPortal database, we found that the alteration frequency of HSP90AB1 varies a lot in different types of cancer. Among all the cancers investigated, the alteration frequency of lung cancer ranked first, reaching about 16%, and amplification was the most common type among mutation, amplification and deep deletion, while deep deletion was less frequent in studied cancers (Figure 6B). What’s more, the survival curves of ACC and BLCA demonstrated that tumor patients with genetic alterations in HSP90AB1 had shorter survival times, and they had lower survive probability typically in stage Ⅲ and Ⅳ (Figure 6C).
Phosphorylation of HSP90AB1 in cancer in the CPTAC database
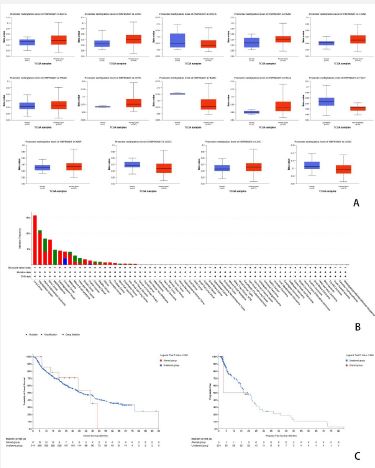

As is shown in (Figure 7). Phosphorylation is a form of protein modification, playing an important role in cell signaling and metabolic regulation. Thus, it’s of great importance to study phosphorylation of HSP90AB1 in different cancers. Nine sets of data showing phosphorylation of HSP90AB1 were obtained from the CPTAC database. By studying these nine sets of data, it’s clear that in glioblastoma multiforme, lower level of HSP90AB1 phosphorylation was observed. In contrast, it was higher in primary tumor than normal in other 8 types of cancers, including Ovarian cancer, Colon cancer, Clear cell RCC, Lung adenocarcinoma, Lung adenocarcinoma, Head and neck squamous carcinoma, Pancreatic adenocarcinoma and Hepatocellular carcinoma. It’s convinced that phosphorylation of HSP90AB1 is higher in most kinds of cancers. In most of the cancers studied, Z-values are more dispersed than in normal tissue, except Colon cancer.


Functional enrichment analysis of HSP90AB1
As is shown in (Figure 8), KEGG and GSEA enrichment analyses were performed to examine HSP90AB1-associated signaling pathways that were differentially activated in cancer. 10% of the differential genes screened were concentrated in the neuroactive ligand-receptor interaction, and the number of genes was 50, and the correlation was high. The selected differential genes were 5% enriched in the signaling receptor activator activity and the receptor ligand activity, respectively, and both of the numbers of genes were 60 (Figure 8A). Through GSEA enrichment analysis, we found that in biological process, HSP90AB1 enriched remarkably in metabolism of amino acids and derivatives and zinc homeostasis. In cellular component, gene enrichment at the top in transport of inorganic cations anions and amino acids oligopeptides and extracellular matrix organization. In KEGG analysis, gene enrichment at the bottom in G Alpha Q signaling events and metabolism of lipids (Figure 8B).
HSP90AB1 expression is associated with immune infiltration
As is shown in (Figure 9), due to the obvious relationship between HSP90AB1 and immune response, we conducted a pan-cancer analysis of the association between HSP90AB1 expression and immune infiltration level based on the TIMER database. We found that most of the 24 kinds of immune cells, including Th2 cells, T helper cells and Tcm, were most positively associated with the HSP90AB1 expression, while some kinds of immune cells, like B cells, Cytotoxic cells, DC, pDC and T cells, were most negatively associated with the HSP90AB1 expression. In a few cells, such as Tem, the correlation between immune cells and HSP90AB1 expression in different types of cancers was not remarkable. Among 33 kinds of cancers, we found that the HSP90AB1 expression negatively correlated with these types in ACC, LUAD, LUSC, SARC, SKCM, STAD and UCS significantly, and the expression of HSP90AB1 correlated with KIRC positively. In conclusion, HSP90AB1 expression negatively correlated with most of the cancer types.
Correlation of HSP90AB1 expression and immune checkpoint Gdlene


As is shown in (Figure 10). Immune checkpoint is one of the main causes of immune tolerance in the process of tumor development. We investigated 24 types of immune checkpoint gene, finding the correlations of HSP90AB1 expression and immune checkpoint gene in 30 kinds of cancers. On the one hand, we found that most of the 24 kinds of immune cells were negatively associated with the HSP90AB1 expression, including CD160, CD244, CTLA4 and LAG3, while some kinds of immune cells, like PVRL2, were most positively associated with the HSP90AB1 expression. Moreover, the correlation of KIR2DL1, KIR2DL3 and HSP90AB1 expression was poor. On the other hand, HSP90AB1 expression negatively correlated with most types significantly, such as COAD, LGG, LUAD, LUSC, OV, PAAD and SKCM, while the expression of HSP90AB1 correlated with ACC and CHOL positively, and the correlation of BLCA, MESO, PCPG and HSP90AB1 expression was poorer than other cancers. Over all, most of the immune checkpoint genes in pancarcinoma were negatively correlated with HSP90AB1 expression.
HSP90AB1 mRNA increased in pancarcinoma tissues
As is shown in (Figure 11). The expression patterns of HSP90AB1protein and mRNA were similar. In addition, we also tested the accuracy of HSP90AB1gene expression levels. Similarly, the use of HPA data showed increased expression of HSP90AB1 protein in pancarcinoma tissues compared to normal tissues.
Because of its high morbidity and mortality, cancer has turned out to be the major factors that seriously threaten human health. Acting in accordance with the survey, the three most common cancers in the world are breast, lung and colon, while the cancers with the highest mortality are lung cancer, stomach cancer and liver cancer [11]. Targeting on the highly proliferating mutated tumor cells, methods such as surgery, radiation, chemotherapy and immunotherapy, have been investigated [12]. Early detection and effective treatment are important conditions for the prognosis of cancer patients. As a new genetic analysis method, pan-cancer analysis is of great significance for cancer prevention design and personalized treatment strategy. In recent years, more and more research has focused on genome-wide pan-cancer analysis, which has significant implications for the early diagnosis of cancer and the identification of sensitive biomarkers [13,14], saving countless cancer patients.
Heat Shock Proteins (HSP) are one of the oldest heat stress proteins in organisms varying between bacteria to mammals. When an organism is exposed to extreme temperatures, it is stimulated by heat to produce this protein, which protects the organism itself. Many heat shock proteins have chaperone activity [15,16]. Most of the HSP genes are not normally expressed, and are usually significantly expressed when the cell is vulnerable to external stimulation. Non-lethal thermal stimulation can strengthen the resistance of organisms to the subsequent thermal stimulation and improve the survival rate of organisms to the lethal thermal stimulation. It has been found that in addition to thermal stimulation, there are countless physical and chemical stimuli that can activate the expression of small molecules of heat shock proteins, such as ultraviolet light, radiation, mechanical damage, acids, oxidants, and so on. It can be seen that small molecular heat shock proteins are essential substances to resist external adverse stimuli [17-19]. HSP can bind to many protein molecules and perform a variety of physiological functions: helping amino acid chains fold into the correct three-dimensional structure, removing damaged amino acid chains that cannot be folded correctly, and escorting protein molecules to find their target molecules so as not to be interfered with by other molecules. Heat shock proteins not only protect proteins that are essential to basic physiological processes, they also break down damaged proteins and recover the raw materials for protein synthesis, allowing the physiological processes inside the cell to run smoothly [20].
Heat shock Response (HSR) is a vital mechanism used by organisms to resist environmental stress and maintain protein homeostasis during life activities [21]. Heat shock can cause structural and functional damage to numerous proteins within the cell, and at this time, the cell initiates the heat shock response to rapidly express heat shock proteins, preventing protein misfolding and aggregation in response to increased temperature and other stress triggered protein homeostasis imbalances [22]. During intense stress-induced heat shock response, the heat shock transcription factor HSP90AB1 undergoes complex activation regulation to rapidly and extensively regulate the transcription of heat shock proteins.
HSP90 is an significant protein that exists in a variety of cell tissues and has a high degree of expression and stability [23]. The structure and function of HSP90 may vary in different organisms. However, in all known higher organisms, the primary function of HSP90 is necessary for cell survival. HSP90 plays an essential role in binding and transporting molecules, maintaining intracellular osmotic pressure and participating in cell apoptosis [24,25].
As a novel biomarker, HSP90AB1 has been widely used in drug screening and biomedical research in recent years. HSP90AB1 (Heat shock protein 90 kDAα, Class B, Member 1), also known as HSP90beta, encodes a member of the heat shock protein 90 family and is an integral part of the heat shock protein family. These proteins play a role in signal transduction, protein folding and degradation, and morphological evolution. This gene encodes a constitutive form of the cytoplasmic 90kDa heat shock protein, which is considered to play a role in apoptosis and inflammation of stomach cells. Variable splicing results in multiple transcript variants [26-28]. The protein transcribed by this gene serves on the HSP family and is a molecular chaperone. Chaperones support proper protein folding and maintain protein stability by constraining to client proteins after exposure to various cellular stresses. Customer proteins belong to various protein families, including kinases, ubiquitous ligases, and transcription factors. The HSP90 protein acts as a dimer and binds the consumer with the help of a co-partner [29]. Chaperone affects many functions, including client binding, ATPase activity, or ATP binding by HSP90, which is necessary for large-scale cellular processes and is therefore critical for cell survival. Based on RNA sequencing data, HSP90AB1 was identified as the most promising target among the other predicted HSPS. In addition, HSP90AB1, which is highly denominated in a variety of cancers, also promotes tumor formation and cancer cell proliferation, migration, and glycolysis, and is associated with poor prognosis [30,31]. As such, they are also targets for innovative treatments for cancer treatment.
Further studies are needed to investigate the molecular mechanisms of HSP90AB1 in pancreatectomy, including how it regulates HSP expression and its role in cell signaling and the tumor microenvironment. In addition, given the potential impact of HSP90AB1 on pan-cancer prognosis, future studies should also evaluate its potential as a therapeutic target and whether intervention strategies targeting HSP90AB1 could provide additional treatment options for cancer patients.
Using data from the TCGA and CPTAC database, we investigated the expression of HSP90AB1 and correlations of HSP90AB1 expression with major pathological stages in a variety of cancer tissues [32] (Figure 1). We observed that protein expression of HSP90AB1 was significantly unregulated in multiple samples compared to normal tissues. And the statistical significance of uncommon cancers and HSP90AB1 expression varied a lot. However, HSP90AB1 expression showed no remarkable change among different cancer stages (Figure 2). And our KEGG and GSEA analyses show that HSP90AB1 is significantly associated with a number of signaling pathways (Figure 8). In biological processes, HSP90AB1 is markedly enriched in amino acid metabolism and zinc homeostasis. In cellular components, genes are enriched in the transport of inorganic cations, anions, amino acids, oligopeptides and at the apex of extracellular matrix tissues. Together, our study provides insights into the application of HSP90AB1 as a potential prognostic biomarker for multiple cancers in an immunooncological context and contributes to the development of targeted therapeutic strategies for HSP90AB1.
Through cBioPortal database, we performed survival association analyses (including OS, DSS and PFI) to determine the association between HSP90AB1 expression levels and prognosis for 33 cancer types (Figures 3-5). Forest plots and survival curves of HSP90AB1 expression on tumor OS, DSS and PFI were observed. OS, DSS and PFI analysis suggested that HSP90AB1 was a risk factor in the patients with most kinds of cancers, while the opposite for a few cancers such as OV. And over all, survival curves showed that patients with higher HSP90AB1 expression had a significantly worse prognosis than those with lower HSP90AB1 expression. This suggests that HSP90AB1 may play a promoting role in the pathological process, and the increased expression of HSP90AB1 may be related to the aggressiveness of the tumor, the ability of metastasis and the resistance to tumor therapy. (Figure 11) showed that the expression of HSP90AB1 messenger RNA (mRNA) was increased in pancreatectomy tissues compared to normal tissues. Therefore, the regulatory mechanisms of HSP90AB1 expression, its unique role in cancer development, and its possibility as a potential therapeutic target are all areas that are necessary to explored in future research.
DNA methylation constitutes one of the earliest discovered and most deeply studied epigenetic regulatory mechanisms. As a relatively stable modification state, DNA methylation plays an important epigenetic mechanism that can be transmitted to newborn offspring DNA during the process of DNA replication [33]. In addition, other mutation types can also have a significant influence on the biological phenotype. Cancer cells are characterized by abnormal DNA methylation (i.e., genome-wide hyperventilation and site-specific hypermethylation) that primarily targets CpG islands in gene expression regulatory elements. Hypermethylation within the promoter region often leads to the silencing or inactivation of tumor suppressor genes in cancer cells [34,35]. The promoter methylation levels of HSP90AB1 in primary tumor were higher than normal in most cancers, which indicated that HSP90AB1 expressed more in most cancers. What’s more, the alteration frequency of HSP90AB1 varies a lot in different types of cancer. These mutations may lead to more active gene expression (Figure 6).
Post-Translational Modification (PTM) is involved in a variety of cellular activities, and phosphorylation is one of the most widely studied PTMS, and alterations in phosphorylation pathways can lead to momentous consequences for disease forms, especially cancer [36]. During protein phosphorylation, protein-protein interactions alter the conformation, activity, localization, and stability of the protein, as well as signal transduction to the next step. Studies have shown that many signaling pathways, which are major players in the cell cycle, are dissimulated in the phosphorylation-dephosphorylation cascade that manifests itself in various types of cancer [37]. In our study, as (Figure 7) showed, HSP90AB1 phosphorylation was higher in primary tumor than normal, which verified the correlation of phosphorylation and cancer.
The Tumor Microenvironment (TME), a complex structure composed of tumor cells, non-malignant cells, blood vessels, extracellular matrix and other substances, is a specialized ecosystem composed of host components designed by tumor cells to influence tumor growth, metastasis, spread, and response to therapy [38]. Diverse types of immune cells play a key regulatory role in TME. Relevant studies have shown that there is a very complex interaction between cancer cells and assorted components of TME, which promotes tumor immune escape, and ultimately leads to tumor proliferation, recurrence and metastasis [39,40]. Although immunotherapy has made some breakthroughs in the treatment of cancer, there are still many challenges, and the study of different targets and biomarkers can help further improve the therapeutic effectiveness of immunotherapy. The function of HSP90AB1 and its effect on the tumor immune microenvironment have not been fully studied. This study revealed the relationship between HSP90AB1 and tumor immune cells, and investigated the immune status of cancer patients by examining HSP90AB1 expression.
Cytotoxic cells are a kind of immune cells, which have the capability to kill infectious pathogens, tumor cells and abnormal cells. By recognizing and binding to specific antigens on receptors, cytotoxic T cells need to be able to release cytotoxins, which in turn induce apoptosis of target cells. There are two main pathways of Cl-mediated tumor cell death, namely granular exocytosis (perforce and granite) and death ligand [41,42]. In our study, it was most negatively associated with the HSP90AB1 expression. CD160, a Glycosylphosphatidylinositol (GPI) -anchored cell surface glycoprotein, is expressed on cytotoxic Natural Killer (NK) cells and T cell subpopulations and exhibits many unique structural and functional features. It is a novel member of the immune superfamily and plays a crucial role in activating NK cell cytotoxicity and cytokine production. The co-stimulatory signals provided by its binding with its specific ligand HVEM molecule play a major role in T cell proliferation, activation, cytokine production and B cell proliferation, activation, secretion of antibodies and other processes. It also regulates the immune system and is associated with certain pathologies, such as cancer [43-45]. Through (Figure 9 and Figure 10), we found that most of the 24 kinds of immune checkpoint genes were negatively associated with the HSP90AB1 expression, especially CD160.
This study comprehensively analyzed the relationship between HSP90AB1 gene expression and 24 altered immune cell subtypes. Notably, our results reveal an interesting phenomenon in which most immunosuppressive and immunostimulatory molecules are inversely correlated with HSP90AB1 expression. Through the TIMER database, we found that among the 24 kinds of immune cells, Th2 cells, T helper cells and Tcm cells were positively correlated with HSP90AB1 expression, while B cells, cytotoxic cells, DC cells, pDC cells and T cells were the most negatively correlated with HSP90AB1 expression. In terms of immune checkpoint genes, most of the 24 immune cells were negatively correlated with HSP90AB1 expression, including CD160, CD244, CTLA4 and LAG3, while limited immune cells such as PVRL2 were positively correlated with HSP90AB1 expression.
Our findings further highlight the indispensable role of HSP90AB1 in immunomodulatory processes in multiple cancers and suggest that HSP90AB1 may modulate the tumor microenvironment by influencing immune cell invasion and function. These findings provide a contemporary perspective on HSP90AB1 as a potential target for cancer immunotherapy. Interventions targeting HSP90AB1 may help regulate the composition of immune cells in the tumor microenvironment, thereby enhancing immune monitoring and inhibiting tumor progression.
However, there are a number limitations to the study. First, the current study was carried out with the basis of data retrieved from an online database and further clinical sample studies are needed to confirm our findings. Secondly, we mainly focus on the bioinformatics analysis of HSP90AB1 expression data. In addition, at present, the peculiar action path of HSP90AB1 is still not comprehensive, and further studies need to be carried out at the molecular level. To understand the role of HSP90AB1 in pericardium, future studies will need to take a multi-dimensional approach, including but not limited to in vitro experiments, animal models, and clinical sample analysis. Objective: To disclose the unique mechanism of HSP90AB1 in immune cell function and tumor immune escape. In addition, exploring the interaction of HSP90AB1 with other immunomodulators and how they work collaboratively to influence the immune microenvironment and therapeutic response of cancer will be key to advancing the development of cancer immunotherapy. Through these studies, we hope that i can provide more accurate and effective treatment strategies for cancer patients and improve patient outcomes and quality of life.
Upregulation of HSP90AB1 expression corresponds to poor prognosis in patients and is associated with infiltration levels of CD160, CD244, CTLA4, B cells, Cytotoxic cells, DC, pDC, T cells, and LAG3 in different cancers. Increased HSP90AB1 phosphorylation and decreased HSP90AB1 methylation have been pointed out in many types of cancer. HSP90AB1 expression was substantially correlated with the expression of immune checkpoint markers. Future prospective and experimental studies of HSP90AB1 expression and immune cell infiltration in diverse cancer populations may provide additional insights into tumor mechanisms and the development of therapeutic strategies targeting HSP90AB1 to improve the therapeutic efficacy of immunotherapy.
Ethical approval: All data were obtained from public databases.
Consent to participate: All the authors agreed to participate in the study.
Consent to publish: All the authors agreed to publish the experimental study.
Authors contributions: All members of this article participated in the first draft of the article writing, Li,Geng,Wangresponsible for most of the writing of the article. Liu responsible for the experimental data visualization.
Funding: No funding was available for this study.
Competing interests: No conflict of interest.
Availability of data and materials: Data are available upon reasonable request.
