1The First Affiliated Hospital, Harbin Medical University, Harbin, 150076, China.
2College of Veterinary Medicine, Northeast Agricultural University, Harbin, 150030, China.
Shichao Liu
Tel: 18545078137;
Email: LSC10251025@163.com
Received : Oct 28, 2024 Accepted : Nov 14, 2024 Published : Nov 21, 2024 Archived : www.meddiscoveries.org
Zinc is essential for cellular processes and has been implicated in the development of various malignancies. The protein SLC39A10, which facilitates zinc transport, is suspected to contribute to tumor growth in multiple types of cancer. Our research delves into the function of SLC39A10 in Hepatocellular Carcinoma (HCC), particularly its impact on immune system reactions and its significance as a predictor of patient outcomes. By examining data from The Cancer Genome Atlas, GEO, and HPA, we observed that SLC39A10 levels were notably higher in HCC tissues compared to non-cancerous ones. This increase in SLC39A10 expression was associated with more severe stages and grades of cancer. Individuals with elevated SLC39A10 levels tended to have a less favorable prognosis in terms of overall survival. We mapped a protein interaction network involving SLC39A10 and explored its potential links to the presence of immune cells within tumors. This analysis was conducted using Gene Ontology, We employed Gene Set Enrichment Analysis (GSEA) and single-sample GSEA to delve into the relationship between SLC39A10 gene expression and immune cell infiltration. By leveraging databases like the Gene Expression Profiling Interactive Analysis and the Tumor Immune Estimation Resource, we examined how SLC39A10 levels correspond with immune cell infiltration markers. The findings from our study suggest that SLC39A10 may play a pivotal role in the progression of Hepatocellular Carcinoma (HCC) and in the tumor’s capacity to elude immune responses, highlighting its potential as a prognostic biomarker for patient outcomes. This revised version aims to provide a fresh perspective on the original text while preserving the scientific findings and conclusions.
Keywords: SLC39A10; ZIP10; Hepatocellular carcinoma; Zinc ion transporter.
Zinc (Zn) is gaining recognition in biological and medical research for its role in cellular homeostasis [1]. It plays a pivotal part in gene expression, immune responses, and cell cycle regulation, acting as a cofactor for numerous enzymes [2]. Zinc’s involvement in cellular processes, including apoptosis and proliferation, is closely tied to its influence on cancer development [3]. The equilibrium of zinc within cells is critical for health, and it significantly impacts cell survival and death [4]. Its role in cell signaling, proliferation, differentiation, and response to oxidative stress suggests zinc’s dualistic potential in tumorigenesis.
Recent advances in understanding zinc ion transport, particularly the function of the solute carrier family 39 (SLC39/ZIP), have highlighted SLC39A10 (ZIP10) due to its increased presence in various cancers and its contribution to tumor growth [5-7]. However, the specific mechanisms of SLC39A10 in Hepatocellular Carcinoma (HCC) and its implications for tumor immunity and patient prognosis are not yet fully elucidated, warranting further investigation.
Hepatocellular Carcinoma (HCC) ranks as the third leading cause of cancer-related deaths worldwide [8]. Its high recurrence and metastasis rates, along with the challenges of treating advanced stages, make improving treatment outcomes a critical research area [9]. Although targeted therapies like Sorafenib and Lenvatinib have shown promise, they are not without side effects and have limitations in their application [10,11]. Identifying novel therapeutic targets and prognostic markers is vital for enhancing patient outcomes in HCC.
Our research endeavored to investigate the expression levels of SLC39A10 in Hepatocellular Carcinoma (HCC) and its association with the progression and prognosis of the disease. By harnessing bioinformatics tools including The Cancer Genome Atlas (TCGA), Gene Expression Omnibus (GEO), and Human Protein Atlas (HPA), we evaluated the expression patterns of SLC39A10. Through the application of multivariate analysis and Kaplan-Meier survival curves, we established a link between SLC39A10 expression and clinical parameters, as well as Overall Survival (OS). Furthermore, we mapped out protein-protein interaction networks to uncover possible genes or proteins that interact with SLC39A10. We also utilized Gene Ontology, GSEA, and single-sample GSEA to scrutinize the relationship between SLC39A10 expression levels and the presence of immune cells in HCC. To deepen our understanding, we analyzed the correlation between SLC39A10 expression and markers of immune cell infiltration, using resources such as the Gene Expression Profiling Interactive Analysis and the Tumor Immune Estimation Resource. This comprehensive approach allowed us to explore the multifaceted role of SLC39A10 in HCC, from its potential as a prognostic indicator to its influence on the tumor’s interaction with the immune system.
These methodologies are expected to provide a comprehensive understanding of SLC39A10’s role in HCC, its impact on the tumor immune microenvironment, and its potential as a prognostic biomarker. The insights gained could offer new molecular targets for HCC and aid in the development of diagnostic and therapeutic strategies, ultimately benefiting clinical management for HCC patients (Figure S1).

Data source: The Cancer Genome Atlas (TCGA) (https://portal.gdc.cancer.gov) is a comprehensive and publicly accessible resource that offers a vast array of clinical and molecular data across various cancer types, including 33 different types of cancer. It serves as a valuable platform for researchers and scholars conducting cancer-related research.
In the context of Hepatocellular Carcinoma (HCC), the TCGA database provides clinical data along with high-throughput RNA sequencing (RNA-Seq) information for patients. The transcriptional expression levels are determined using the per kilobase of transcript per million mapped reads (FPKM) method as implemented in HTSeq. Furthermore, the RNA-Seq gene expression levels, originally quantified as FPKM, are converted into Transcripts Per Million (TPM) for 539 HCC patients to facilitate further analysis.
The GEO and HPA databases
The Gene Expression Omnibus (GEO) is a prominent public repository that collects and stores gene expression data derived from high-throughput experimental studies. It plays a pivotal role in genomic research and is overseen by the National Center for Biotechnology Information (NCBI). Researchers can access this valuable resource at the NCBI GEO website to retrieve a wealth of information for their studies. In addition to GEO, the Human Protein Atlas (HPA) serves as an essential companion database, providing an in-depth dataset on the expression and distribution of human proteins across various tissues and cell types. With its coverage of 44 normal tissues and 20 prevalent cancer types, the HPA offers a comprehensive perspective on protein expression in both healthy and disease states. Moreover, the HPA includes immunohistochemistry data for proteins in both tumor and normal tissue samples, which is particularly useful for deciphering the roles of proteins within different biological contexts.
Clinical statistical analysis of prognosis, model development, and assessment
The Xiantao platform, as described, offers a comprehensive suite of tools for bioinformatics analysis, including the application of clinical data from TCGA to assess patient outcomes in Hepatocellular Carcinoma (HCC). On the Xiantao platform, we conducted a thorough analysis to understand the role of SLC39A10 gene expression in Hepatocellular Carcinoma (HCC). We used statistical methods such as Cox regression and Kaplan-Meier survival analysis to evaluate key survival metrics for HCC patients, including Overall Survival (OS), Disease-Specific Survival (DSS), and Progression-Free Survival (PFS). Patients were categorized into low and high expression groups based on the median values of SLC39A10 gene expression. To delve into how SLC39A10 expression relates to clinical characteristics, we employed the Wilcoxon signed rank sum test and logistic regression analysis. Additionally, a multivariate Cox regression model was utilized to evaluate the influence of SLC39A10 expression on survival probabilities, along with other clinical variables. We considered a p-value less than 0.05 as our threshold for statistical significance.
The Cox regression model was integrated with other independent prognostic factors to predict 1-year, 3-year, and 5-year survival rates. To ensure the reliability of our predictions, a calibration curve was generated, allowing us to compare the predicted survival rates with the actual rates observed in the study population. This approach provided a robust framework for assessing the clinical implications of SLC39A10 expression in HCC. Ideally, the curve should align with the 45-degree line, indicating precise predictive accuracy. This approach allows for a thorough understanding of the clinical implications of SLC39A10 gene expression in HCC and its potential as a prognostic indicator.
Comprehensive protein-protein interaction analysis
The STRING database (https://string-db.org/) is ideal for analyzing data. It offers a comprehensive collection of protein-protein interaction (PPI) data. By inputting SLC39A10 expression data into STRING, we obtained PPI network information, using a confidence score above 0.7 as our threshold for significance.
Enrichment analysis
We performed a Gene Ontology (GO) enrichment analysis to uncover the biological roles of the SLC39A10 gene. Utilizing the cluster Profiler package in R, version 3.6.3, we sorted the differentially expressed genes into CC, MF, and BP GO categories. We fine-tuned our analysis by setting specific parameters: the enrichment coefficient was set higher than 1.5, a minimum count of 3 was required, and the p-value threshold was placed at less than 0.01. Furthermore, gene set enrichment analysis (GSEA) was conducted to sequence the genome and spot pathways closely linked to SLC39A10 gene expression. This process was repeated a thousand times for each analysis. In GSEA, statistical significance was determined with an adjusted p-value under 0.05 and a False Discovery Rate (FDR) under 0.25. The results of the enrichment analysis were evaluated using Normalized Enrichment Scores (NES) and adjusted p-values, indicating how strongly the gene set was associated with a specific biological theme. For GSEA, we used a cluster analyzer tool for both conducting the analysis and visualizing the outcomes. This systematic approach provides in-depth insights into the biological functions of SLC39A10 and its potential role in a range of cellular pathways, which is vital for comprehending its impact on disease progression and therapeutic outcomes.
Analysis of infiltration of immune cells
We applied the single-sample gene set enrichment analysis (ssGSEA) to evaluate immune cell infiltration in tumors, examining 24 different immune cell types. The Spearman correlation algorithm was used to measure the correlation between SLC39A10 gene expression levels and immune cell infiltration. This method enabled us to assess the differences in immune cell infiltration between groups with low versus high SLC39A10 expression and to determine the strength of the association for each of the 24 immune cell types. Leveraging the findings from the immune infiltration analysis, we conducted further analysis on the Xiantao Tool module using the Spearman correlation and Wilcoxon signed-rank test to explore the link between SLC39A10 gene expression and immune cell infiltration. This helped elucidate the relationship between immune cell infiltration levels and SLC39A10 gene expression subset values. Statistical significance was set at a p-value threshold of less than 0.05, with additional significance levels marked as *p< 0.05, **p< 0.01, ***p< 0.001, and ****p< 0.0001.
Gene correlation analysis
GEPIA (http://gepia.cancer-pku.cn/index.html) is a powerful online tool that provides a wide-ranging dataset, including 9,736 types of cancer and 8,587 normal tissue samples from TCGA and GTEx. It offers an extensive gene expression data repository, with 60,498 gene classes and 198,619 isomer classes at our disposal for analysis. We utilized GEPIA to investigate how SLC39A10 gene expression correlates with various immune cell markers. Within our analysis, we charted SLC39A10 gene expression on the X-axis and the expression levels of other pertinent genes on the Y-axis. Additionally, we used TIMER (http://cistrome.org/TIMER/) data to evaluate the relationship between gene expression patterns and SLC39A10. We set the bar for statistical significance at a p-value less than 0.05, with finer grades of significance noted as *p< 0.05, **p< 0.01, ***p< 0.001, and ****p< 0.0001.
The gene expression of SLC39A10 was elevated in hepatocellular carcinoma compared to normal tissue
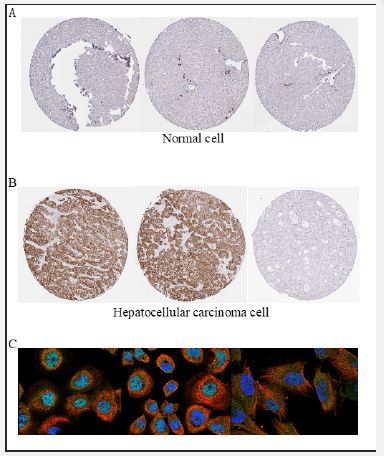
To ascertain whether elevated SLC39A10 gene expression is a common characteristic in cancers, a pan-cancer analysis was conducted. We compared SLC39A10 gene expression levels in HCC tumor samples with those in adjacent healthy tissues within the TCGA dataset (Figure 1A). Our study discovered significantly elevated levels of SLC39A10 mRNA in HCC tumor samples when compared to normal tissues (p< 0.001) (Figure 1B). Further analysis compared SLC39A10 expression in normal tissues from the GTEx database, para-HCC tissues, and HCC tissues, confirming upregulation of SLC39A10 in HCC (p< 0.001) (Figure 1C). There was also notable deregulation of SLC39A10 in HCC compared to adjacent non-cancerous samples (p< 0.001) (Figure 1D). An ROC curve was generated to evaluate the diagnostic value of SLC39A10 expression, resulting in an AUC of 0.694, confidence interval 0.694-0.740, indicating a significant diagnostic potential for HCC (Figure 1E). In alignment with mRNA levels, SLC39A10 protein expression was higher in HCC tissues than in normal tissues (Figure 1F), suggesting a match between protein and mRNA expression in our dataset. Further assessment of SLC39A10’s diagnostic accuracy was carried out (Figures 2A,B). HPA data showed increased SLC39A10 protein expression in HCC tissues compared to normal tissues (Figure 2C), underscoring the possibility of SLC39A10 serving as a HCC biomarker.
The relationship between SLC39A10 expression levels and various clinical parameters was examined
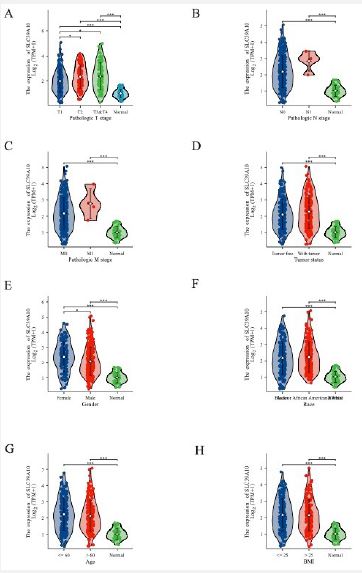
To determine the clinical significance of SLC39A10 expression in HCC, we stratified tumor samples into low and high expression groups based on SLC39A10 expression levels, utilizing the Z-score criterion. The Kruskal-Wallis and Wilcoxon sign-Rank tests were used to evaluate the correlation between SLC39A10 expression and different clinical parameters. We found significant associations between higher SLC39A10 expression and advanced T, N, M categories, tumor status, primary treatment outcomes (PD), and OS events (death) (p< 0.05, Figure 3A-H). Furthermore, univariate and multivariate analyses of HR and 95% CI for various patient characteristics with HCC were performed (Table 1). Univariate analysis showed a substantial link between SLC39A10 expression and several clinical parameters, including Pathologic T stage (OR=1.525, 95% CI 1.013-2.296, p=0.043), Pathologic stage (OR=1.671, 95% CI 1.028-2.716), Tumor status (OR=1.362, 95% CI 0.894-2.076), race (OR=1.088, 95% CI 0.719-1.648, p=0.038), residual tumor (OR=2.885, 95% CI 1.006-8.278, p=0.049), and histologic grade (OR=2.412, 95% CI 1.561-3.726, p< 0.001) (Table 2). These results indicate that SLC39A10 expression is correlated with the clinical features of HCC, emphasizing its potential as a prognostic biomarker. The correlation with clinical parameters such as tumor stage, treatment outcomes, and survival events highlights the importance of SLC39A10 in HCC progression and prognosis.
| Characteristic | Total (N) | HR (95% CI) Univariate analysis | P Value | HR (95% CI) Multivariate analysis | P Value |
|---|---|---|---|---|---|
| Pathologic T stage | 370 | ||||
| TI&T2 | 277 | ||||
| T3&T4 | 93 | 2.598(1.826-3.697) | < 0.001 | 1.930(0.261 14.290) | 0.520 |
| Pathologic N stage | 258 | ||||
| NO | 254 | ||||
| N1 | 4 | 2.029(0.497 8.281) | 0.324 | ||
| Pathologic M stage | 272 | ||||
| M0 | 268 | ||||
| MI | 4 | 4.077(1.281 12.973) | 0.017 | 0.973(0.230-4.121) | 0.971 |
| Pathologic stage | 349 | ||||
| Stage I & Stage II | 259 | ||||
| Stage III & Stage IV | 90 | 2.504(1.727 3.631) | < 0.001 | 1.297(0.176 9.538) | 0.798 |
| Tumor status | 354 | ||||
| Tumor free | 202 | ||||
| With tumor | 152 | 2.317(1.590 - 3.376) | < 0.001 | 1,847(1.152 2.962) | 0.011 |
| Gender | 373 | ||||
| Female | 121 | ||||
| Male | 252 | 0.793(0.557 1.130) | 0.200 | ||
| Race | 361 | ||||
| Asian | 159 | ||||
| White & Black or African | 202 | 1.341(0.926 1.942) | 0.121 | ||
| American | |||||
| Age | 373 | ||||
| <= 60 | 177 | ||||
| > 60 | 196 | 1.205(0.850 1.708) | 0.295 | ||
| Weight | 345 | ||||
| <= 70 | 184 | ||||
| > 70 | 161 | 0.941(0.657 1.346) | 0.738 | ||
| Histological type | 373 | ||||
| Hepatocellular carcinoma | 363 | ||||
| Hepatocholangiocarcinoma (mixed) & Fibrolamellar | 10 | 0.439(0.061 3.145) | 0.412 | ||
| carcinoma | |||||
| Residual tumor | 344 | ||||
| R0 | 326 | ||||
| R1&R2 | 18 | 1.604(0.812 3.169) | 0.174 | ||
| Histologic grade | 368 | ||||
| G1&G2 | 233 | ||||
| G3&G4 | 135 | 1.091(0.761 1.564) | 0.636 | ||
| SLC39A10 | 373 | ||||
| Low | 187 | ||||
| High | 186 | 1.812(1.274 2.577) | < 0.001 | 2.012(1.249 3.243) | 0.004 |
| Characteristics | Total (N) | OR (95% CI) | P value |
|---|---|---|---|
| Pathologic T stage (T2&T3&T4 vs. T1) | 371 | 1.525(1.013-2.296) | 0.043 |
| Pathologic N stage (N1 vs. NO) | 258 | 3.048(0.313 29.689) | 0.337 |
| Pathologic M stage (M1 vs. M0) | 272 | 3.045(0.313 29.642) | 0.338 |
| Pathologic stage (Stage III &Stage IVvs. Stage I&Stage II) | 350 | 1.671(1.028 2.716) | 0.038 |
| Tumor status (With tumor vs. Tumor free) | 355 | 1.362(0.894 2.076) | 0.150 |
| Gender (Male vs. Female) | 374 | 0.440(0.281 0.687) | < 0.001 |
| Race (White&Black or African American vs. Asian) | 362 | 1.088(0.719 1.648) | 0.690 |
| Age (> 60 vs. <= 60) | 373 | 0.781(0.520-1.174) | 0.234 |
| Histological type (Hepatocholangiocarcinoma (mixed) & Fibrolamellar carcinoma vs. Hepatocellular carcinoma) | 374 | 2.385(0.607 9.366) | 0.213 |
| Residual tumor (R1&R2 vs. R0) | 345 | 2.885(1.006 8.278) | 0.049 |
| Histologic grade (G3&G4 vs. G1&G2) | 369 | 2.412(1.561 3.726) | < 0.001 |
| Weight (>70vs. <= 70) | 346 | 0.846(0.554 1.291) | 0.439 |
Bold values are used to highlight statistical significance, and P values are less than 0.05.
Correlation of SLC39A10 expression in HCC prognosis
The TCGA database analysis uncovered a significant link between SLC39A10 expression levels and critical prognostic indicators like Overall Survival (OS), Disease-Specific Survival (DSS), and Progression-Free Survival (PFS). Patients exhibiting higher SLC39A10 expression were associated with worse OS (HR=0.51, 95% CI 0.37-0.69, p< 0.001, Figure 4A), DSS (HR=0.40, 95% CI 0.27-0.60, p< 0.001, Figure 4B), and PFS (HR=0.57, 95% CI 0.41-0.79, p< 0.001, Figure 4C). This correlation suggests that elevated SLC39A10 expression is tied to a higher risk score, while lower scores correspond to reduced SLC39A10 expression. Moreover, the study investigated the connection between SLC39A10 expression and certain patient subgroups, finding that those in later tumor stages (T3-T4) had increased SLC39A10 expression (HR=0.58, 95% CI 0.39-0.86, p=0.007), as did patients with pathological grade III-IV (HR=0.63, 95% CI 0.44-0.91, p=0.015) and histological grade G3-G4 (HR=0.60, 95% CI 0.42-0.85, p=0.005) (Figure 4D). A risk score model for HCC, including variables like M-stage, pathologic grade, N-stage, histological grade, age, T-stage, and SLC39A10 expression, was formulated to refine clinical prognosis (Figure 4E). The accuracy of these model predictions was gauged using calibration plots (Figure 4F). With this model, the 3-year and 5-year survival rates could be forecasted more precisely, considering SLC39A10 expression.In conclusion, SLC39A10 expression correlates with the prognosis of HCC patients, indicating its potential as a biomarker for risk assessment and personalized therapeutic approaches.
Individuals with elevated SLC39A10 expression levels exhibited poorer prognosis indicators than those with lower SLC39A10 expression, such as reduced Overall Survival (OS) (A), shorter Progression-Free Survival (PFS) (B), and diminished Disease-Specific Survival (DSS) (C) (both log-rank p< 0.001). (D) The prognosis varied based on SLC39A10 expression across various clinical features for OS. (E) A clinical features-based nomogram, incorporating SLC39A10 expression, was developed for multivariate analysis. (F) The calibration plot illustrates the accuracy of the model’s predictions, as determined by multi-factor Cox regression analysis.
Build PPI network
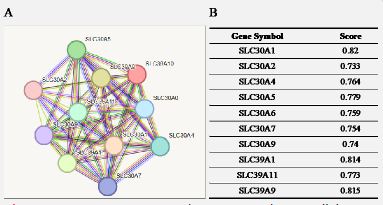
Understanding functional interactions between proteins is crucial to further exploring the molecular basis and metabolic processes involved in cancer. By using the STRING program, the SLC39A10PPI network was analyzed and the proteins interacting with SLC39A10 were identified. Figure 5 shows the top eleven proteins and their associated gene names, scores, and annotations. These include SLC30A1, SLC30A2, SLC30A4, SLC30A5, SLC30A6, SLC30A7, SLC30A9, SLC39A1, SLC39A11, SLC39A9.
Expression of SLC39A10 gene and whole gene expression pattern
To further explore the biological role of the SLC39A10 gene in HCC, a comprehensive analysis of its gene expression was performed. A significant correlation was identified between SLC39A10 expression and both down-modulated and up-modulated genes, with a log fold change greater than 1 and an adjusted p-value less than 0.05 (Figure 6A). This analysis helps to identify genes that may be co-regulated with or influenced by SLC39A10. A heatmap was created to display the top 20 genes with the most significant expression changes, with an absolute log fold change greater than 2 and an adjusted p-value less than 0.01 (Figure 6B, Table 3). These genes are considered potential candidates for further investigation into their roles in HCC biology. To understand the biological functions associated with SLC39A10, a Gene Ontology (GO) enrichment analysis was conducted. The results showed that SLC39A10 is mainly linked to cellular components such as the apical plasma membrane and the apical part of the cell, as well as molecular functions like ion channel complex formation. The biological processes involving SLC39A10 include pattern specification, embryonic organ morphogenesis, and regionalization. At the molecular function level, SLC39A10 is involved in metal ion transmembrane transport, channel activity, and ion channel activity. Furthermore, a Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis was performed to identify pathways related to SLC39A10 expression. This analysis revealed close associations between the SLC39A10 gene and pathways such as mineral absorption, neuroactive ligand-receptor interaction, and the metabolism of xenobiotics by cytochrome P450. These findings offer valuable insights into the potential roles of SLC39A10 in HCC, suggesting its involvement in critical biological processes and pathways. Understanding these relationships can guide future research directions, such as exploring the therapeutic potential of targeting SLC39A10 or related pathways in HCC.

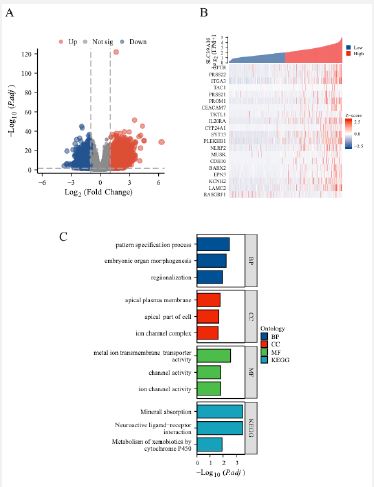
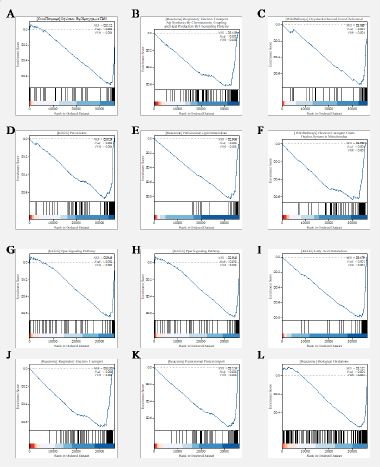
SLC39A10 gene expression of GSEA
GSEA was conducted to uncover the biological pathways and functions that are associated with varying levels of SLC39A10 gene expression. The analysis was guided by the Normalized Enrichment Score (NES), which helped to identify the most relevant pathways in relation to SLC39A10 expression (Figure 7). The pathways that showed significant association with high SLC39A10 gene expression included:
| Gene Name | ID | p-value | p. adjust |
|---|---|---|---|
| CFTR | ENSG00000001626.16 | 3.44e-13 | 7.91e-12 |
| PRSS22 | ENSG00000005001.10 | 2.97e-29 | 1.83e-26 |
| ITGA3 | ENSG00000005884.18 | 1.49e-38 | 6.65e-35 |
| PRSS21 | ENSG00000007038.11 | 1.77e-15 | 6.66e-14 |
| PROM1 | ENSG00000007062.12 | 1.58e-27 | 7.30e-25 |
| CEACAM7 | ENSG00000007306.15 | 2.50e-29 | 1.65e-26 |
| TKTL1 | ENSG00000007350.17 | 2.04e-14 | 6.16e-13 |
| IL20RA | ENSG00000016402.13 | 2.31e-09 | 2.49e-08 |
| CYP24A1 | ENSG00000019186.10 | 5.13e-11 | 7.68e-10 |
| SYT13 | ENSG00000019505.8 | 9.19e-35 | 1.93e-31 |
| PLEKHB1 | ENSG00000021300.14 | 1.49e-28 | 8.19e-26 |
| NLRP2 | ENSG00000022556.16 | 1.79e-28 | 9.64e-26 |
| MUSK | ENSG00000030304.14 | 2.16e-12 | 4.22e-11 |
| CDH10 | ENSG00000040731.10 | 2.19e-10 | 2.87e-09 |
| BARX2 | ENSG00000043039.7 | 6.89e-21 | 8.05e-19 |
| EPN3 | ENSG00000049283.18 | 1.81e-23 | 3.47e-21 |
| KCNH2 | ENSG00000055118.16 | 1.83E-23 | 3.47e-21 |
| LAMC2 | ENSG00000058085.15 | 1.71e-30 | 1.36e-27 |
| RASGRF1 | ENSG00000058335.16 | 3.45e-24 | 7.60e-22 |
Oxidation by Cytochrome P450 (WikiPathways).
Respiratory Electron Transport, ATP Synthesis by Chemiosmotic Coupling, and Heat Production by Uncoupling Proteins (Reactome).
Oxysterols Derived from Cholesterol (WikiPathways).
Peroxisome (KEGG).
Peroxisomal Lipid Metabolism (Reactome).
Cholesterol Metabolism (KEGG).
Electron Transport Chain Oxphos System in Mitochondria (WikiPathways).
PPAR Signaling Pathway (KEGG).
Phase I Functionalization of Compounds (Reactome).
Fatty Acid Metabolism (KEGG).
Respiratory Electron Transport (Reactome).
Peroxisomal Protein Import (Reactome).
Biological Oxidations (Reactome).
These findings suggest that high SLC39A10 gene expression in HCC is associated with a range of biological processes, including lipid metabolism, oxidative stress, and cellular respiration. The involvement of these pathways may provide insights into the role of SLC39A10 in the pathophysiology of HCC and could potentially reveal new therapeutic targets. It’s important to note that while GSEA can suggest biologically relevant pathways, the results should be interpreted with caution and further validated through experimental approaches. The identified pathways could be explored in the context of HCC development and progression, and their association with SLC39A10 could provide a foundation for developing targeted therapies or prognostic markers.
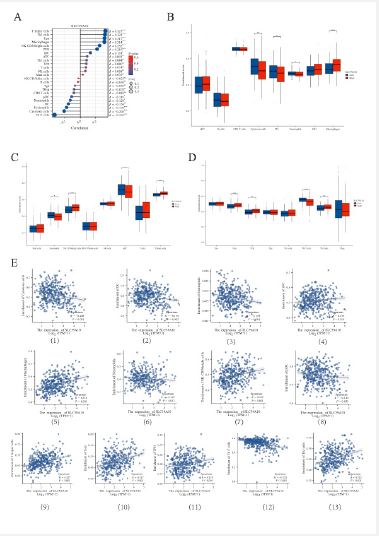
Correlation between SLC39A10 gene expression levels and the presence of immune cells infiltrating tumors
Our study delved into the relationship between SLC39A10 gene expression and the infiltration of 24 distinct immune cell subtypes in Hepatocellular Carcinoma (HCC). We found a robust positive link between SLC39A10 expression and the presence of T helper cells, Th2 cells, Tem cells, and macrophages, while a marked negative correlation was seen with Th17 cells, cytotoxic cells, and eosinophils (Figure 8A, E1-13). These findings align with prior research indicating that elevated ZIP10 levels, crucial for zinc homeostasis, could lead to a reduction in Th17 cells, thereby affecting the tumor’s immune microenvironment. Further investigation showed that the infiltration of immune cells, especially iDCs, macrophages, cytotoxic cells, NK CD56bright cells, pDCs, and T helper cells, notably influenced SLC39A10 gene expression levels (Figure 8B-D). To appraise the impact of SLC39A10 on immune cell infiltration in HCC more comprehensively, we used data from TIMER and GEPIA databases, establishing connections between SLC39A10 and diverse immune markers indicative of various immune cell populations, including DCs, NK cells, M1/M2 macrophages, neutrophils, TAMs, B cells, monocytes, general T cells, and CD8+ T cells (Table S1). The study also examined various functional T cell subtypes like Tregs, depleted T cells, Th1, Th2, Th9, Th17, Th22, and Tfh, revealing that the expression of numerous immune-set markers from different DC types, M1/M2 macrophages, TAMs, and T cells correlated significantly with SLC39A10 gene expression levels in HCC. These results highlight the possible role of SLC39A10 in the formation of the tumor immune microenvironment in HCC, suggesting that targeting SLC39A10 could be a valuable strategy for regulating immune cell infiltration and function. Additional research is required to clarify the mechanisms through which SLC39A10 directs immune cell behavior and to assess its potential as a therapeutic target in HCC.
Hepatocellular Carcinoma (HCC) is among the most common causes of cancer-related mortality globally, underscoring the urgency to understand its molecular underpinnings and develop effective treatments [12,13]. The advent of precision medicine has opened new avenues for targeted therapies, and the SLC39A10 gene has emerged as a promising molecular marker in this context. Our study’s construction of a prognostic model using SLC39A10 gene expression levels aims to shed light on its role in HCC prognosis and equip clinicians with a tool for more precise patient outcome predictions. This could help in developing personalized treatment strategies, potentially enhancing survival rates and the quality of life for HCC patients. By employing both univariate and multivariate Cox regression analyses, we’ve confirmed the independent prognostic significance of SLC39A10. These findings suggest that targeted therapies against SLC39A10 could be a viable strategy to enhance treatment efficacy in HCC patients. The implications of these results are significant for the field of oncology, as they point towards personalized medicine approaches that could lead to more effective treatments with fewer side effects. Future research directions could include:
Mechanistic studies: Investigating how SLC39A10 contributes to HCC progression at the molecular level 2.
Therapeutic development: Designing and testing drugs that target SLC39A10 in preclinical models.
Clinical trials: Conducting trials to evaluate the efficacy of SLC39A10-targeted therapies in patients.
Biomarker validation: Further validating SLC39A10’s prognostic value in larger, diverse patient populations. The potential of SLC39A10 as a therapeutic target and biomarker represents an exciting development in HCC research and could lead to significant advancements in treatment options for patients.
Zinc is an essential trace element with a wide range of biological functions, extending beyond its role as a cofactor. Within the cell, zinc ions (Zn2+) are carefully regulated to maintain cellular function and homeostasis. Zinc homeostasis is crucial for cell survival and plays a significant role. Zinc is a vital micronutrient that plays numerous biological roles, not just as a cofactor. Intracellularly, the levels of zinc ions (Zn2+) are meticulously controlled to sustain cell function and equilibrium [14-16]. Preserving zinc homeostasis is critical for cell vitality and significantly impacts an organism’s health [17-19]. It is especially essential for processes like cell signaling, proliferation, and differentiation, all of which are necessary for maintaining healthy tissue function and balance of an organism [20]. It is particularly important in processes such as cell signaling, cell proliferation, and differentiation, which are vital for proper tissue function and homeostasis.
Zinc also has a significant impact on the immune system [21,22]. Deficiencies in zinc have been linked to decreased immune function, leading to a reduced ability to fight off pathogens [23,14]. Moreover, zinc is essential in regulating the inflammatory response by influencing cytokine production and immune cell activity [21,24]. Sufficient zinc consumption is essential for preserving immune system health and avoiding infections.
In pathological conditions, disruptions in zinc balance can result in diverse pathologies [25]. In certain cancers, high levels of zinc are linked to heightened proliferation and virulence of cancer cells [26]. Conversely, zinc deficiency can degrade the functionality of immune cells in the tumor microenvironment, influencing cancer progression and spread [27]. Therefore, regulating zinc homeostasis could provide novel strategies for cancer treatment, particularly for tumors such as Hepatocellular Carcinoma (HCC).
Grasping the complex interplay between zinc homeostasis and cancer is key to devising novel therapeutic strategies. In Hepatocellular Carcinoma (HCC), focusing on zinc transporters such as SLC39A10 might hinder cancer cell development and survival, offering a potential therapeutic target. However, the link between zinc and cancer is intricate, necessitating further research to comprehend this relationship fully and to devise effective treatments.
Zinc ion (Zn2+) homeostasis is essential for keeping cell function and overall health in organisms. Disruptions in this balance can precipitate numerous diseases. Excessive intracellular zinc accumulation can inhibit cell proliferation and induce cell death by triggering apoptosis. The effect of zinc on mitochondria can lead to the release of cytochrome c, thereby activating the caspase cascade, a key regulatory mechanism for apoptosis [28,29].
Intracellular zinc balance is regulated by zinc transporters, including the SLC39 and SLC30 families. SLC39 family proteins, including SLC39A10, are in charge of zinc uptake from extracellular fluid or intracellular vesicles, raising zinc levels in the cytoplasm. Conversely, SLC30 family proteins move zinc out of the cell to extracellular spaces or intracellular vesicles, lowering cytoplasmic zinc levels [30]. This regulated system of zinc import and export ensures the appropriate distribution and concentration of zinc ions within the cell, which is vital for cell survival and function.
The abnormal expression of SLC39A10 has been a significant focus in cancer research. Studies have shown that uncontrolled expression of SLC39A10 is associated with the progression of various cancers, such as breast cancer and gastric adenocarcinoma [5,31]. In breast cancer, increased levels of SLC39A10 can enhance the aggressiveness and metastatic capacity of tumor cells by promoting zinc ion influx [5,31,32]. In gastric adenocarcinoma, SLC39A10 might activate CK2 by increasing the availability of zinc ions, a kinase implicated in several cancers, with elevated activity linking to increased tumor cell proliferation and survival [5,31].
These findings underscore the significance of understanding the role of zinc transporters in cancer development. The link between SLC39A10’s abnormal expression and cancer progression suggests possible therapeutic approaches. Targeting zinc transporters could interrupt zinc-dependent processes that support cancer cell growth and dissemination, presenting a new direction for cancer treatment.
Although the role of SLC39A10 in various cancers is relatively well-understood, its specific mechanism in HCC is still being explored [6,33]. Our comprehensive analysis has brought to light the up-regulation of SLC39A10 expression in HCC. Importantly, this increase in expression correlates with poorer patient prognosis, suggesting that SLC39A10 may significantly contribute to HCC’s development and progression. It is hypothesized that SLC39A10 might be influencing the behavior of tumor cells in HCC by disrupting zinc ion homeostasis and modulating intracellular signaling pathways.
Further investigation is necessary to clarify the precise molecular mechanisms of SLC39A10 in HCC, including its control over zinc ion transport and its role in cell signaling and the tumor microenvironment. Given the prognostic significance of SLC39A10 expression levels in HCC, future research should consider SLC39A10 as a potential therapeutic target and evaluate whether targeting SLC39A10 could provide novel treatment strategies for HCC patients.
Analysis of data from the GEO, TCGA, and HPA databases has indicated significantly higher SLC39A10 mRNA expression in HCC samples compared to normal tissues, which correlates with a poorer prognosis in patients with higher SLC39A10 expression.
Our detailed analysis with the STRING database has identified key proteins interacting with SLC39A10 in HCC, including SLC30A5, SLC30A2, and SLC39A9. These proteins, part of the solute carrier superfamily, are crucial for maintaining zinc ion homeostasis within the cell by regulating Zn2+ transmembrane transport. They are vital for preserving intracellular environment stability and cell function [34,35]. While their roles in HCC have been preliminarily revealed, further studies are needed to fully understand their interactions and functional networks in HCC development.
The ROC analysis in our study showed an AUC of 0.694 for SLC39A10 in diagnosing HCC, suggesting its potential as a diagnostic biomarker and a modern molecular target for early diagnosis and treatment of the disease. Additional bioinformatics analysis and predictive models have shown a correlation between increased SLC39A10 expression levels and the progression of clinicopathological features in HCC patients, which may predict a worsening of patient prognosis.
In cancer biology, the expression level of the zinc ion transporter SLC39A10 and its role in immune regulation are increasingly being studied. Previous research has indicated that abnormal SLC39A10 expression affects the activity of NK cells and T cell subsets and is associated with lymph node metastasis of breast cancer, pointing to a significant role for SLC39A10 in the tumor immune microenvironment.
Tregs and Th17 cells are crucial for immune homeostasis, with Tregs maintaining immune tolerance and Th17 cells promoting inflammatory responses [36-38]. The balance between these cells is essential for immune system stability, and in HCC, this balance is closely tied to disease progression and metastasis [39]. Our comprehensive analysis of the relationship between SLC39A10 gene expression and 24 different immune cell subtypes in HCC showed a significant positive correlation with the tumor invasion level of T helper cells, Th2 cells, Tem cells, and macrophages. A particularly strong correlation with Th17 cell infiltration was observed, highlighting the complex role of Th17 cells in the tumor immune microenvironment [40,41].
Our findings underscore the critical role of SLC39A10 in the immunomodulatory process of HCC and suggest that it may regulate the tumor microenvironment by influencing immune cell invasion and function. This perspective on SLC39A10 as a potential target for HCC immunotherapy is significant. Targeting SLC39A10 may help regulate the composition of immune cells in the tumor microenvironment, thereby enhancing immune surveillance and inhibiting tumor progression.
However, the study has limitations. It is based on data from online databases, and further studies with clinical samples are needed for confirmation. Our analysis mainly focuses on SLC39A10 expression data, and a more comprehensive understanding of SLC39A10’s role in HCC requires a multi-dimensional approach, including in vitro experiments, animal models, and clinical sample analysis. This will help reveal the distinct mechanism of SLC39A10 in immune cell function and tumor immune escape, as well as its interaction with other immunomodulators. Such studies are key to advancing HCC immunotherapy and developing more accurate and effective treatment strategies, ultimately improving patient prognosis and quality of life.
Our study uncovered the upregulation of the SLC39A10 gene in HCC, correlating with increased tumor aggression and poorer patient prognosis. As a key zinc transporter, SLC39A10’s disruption may modulate cancer cell behavior, influencing zinc balance and cellular signaling.
SLC39A10’s role in HCC is dualistic, promoting tumor growth and altering the tumor microenvironment by affecting immune cell infiltration, particularly Th17 cells. These findings hint at its significant immunomodulatory function and its potential in tumor immune evasion and therapy resistance.
Furthermore, we identified SLC39A10’s interaction with other zinc transporters, suggesting its broader impact on cellular zinc homeostasis. This research sets the stage for future exploration of SLC39A10’s mechanisms in HCC and its therapeutic potential, aiming to enhance treatment efficacy for HCC patients.
Ethical approval: All data were obtained from public databases.
Consent to participate: All the authors agreed to participate in the study.
Authors’ contributions: All members of this article participated in the first draft of the article writing, Li responsible for most of the writing of the article. Liu responsible for the experimental data visualization.
Consent to publish: All the authors agreed to publish the experimental study.
Funding: No funding was available for this study.
Competing interests: No conflict of interest.
Availability of data and materials: Data are available upon reasonable request.
