1Department of Embryology, Reproductive Biomedicine Research Center, Royan Institute for Reproductive Biomedicine, ACECR, Tehran, Iran.
2Department of Cellular and Molecular Biology, School of Biology, Damghan University, Damghan, Iran.
3Department of Biology, Faculty of Basic Sciences, Yasuj University, Yasuj, Iran.
4Faculty of Biotechnology, Amol University of Special Modern Technologies, Amol, Iran.
Mohammad Taghi Ghorbanian
Tel: +98-9125318732 & +98 2335220418;
Email: Ghorbanian@du.ac.ir
Received : Jun 22, 2024 Accepted : Jul 31, 2024 Published : Aug 07, 2024 Archived : www.meddiscoveries.org
Objective: Many neural and behavioral functions are affected by estrogens, including mood, cognitive function, and learning. The hippocampus is one of the areas of the brain whose function is related to cognition, memory, and learning and is involved in Alzheimer’s disease. The present study aimed to investigate the effects of estrogen hormone changes of the estrous cycle on the neuronal morphology in the mice hippocampus.
Methods and materials: Female adult NMRI mice (6-8 weeks) was used in this study. Adult females were divided into five groups: Proestrus, Estrus, Metestrus, Diestrus, and Ovariectomy groups. The animals were perfused transcardially and the brain was sectioned and stained with Golgi and Cresyl Violet methods and observed by microscope. Hormone level estrogen was measured by ELISA. The neuronal densities in microscopic sections were examined using Image J software.
Results: Our findings suggest that the highest amount of estrogen hormone level in blood plasma was in the proestrus stage (p<0.05). The proestrus stage has greater neuronal density and dendritic complexity in dentate gyrus granule neurons and in CA1 pyramidal neurons compared to other groups. Comparing ovariectomy and other phases of the estrous cycle, the proestrus phase had higher levels of density of neurons and dendritic spine, quantified by Golgi and Cresyl Violet staining (p<0.05).
Conclusion: Neurogenesis is correlated to changes in estrogen hormone levels at different phases of the estrus cycle and it can affect cognition, memory, and learning. It can also be suggested as a therapeutic strategy for Alzheimer’s disease.
Keywords: Hippocampal plasticity; Neurogenesis; Estrogen hormone; Estrous cycle; Golgi staining.
The hippocampus is one of the brain regions whose role is related to cognition, memory, and learning, and it is the public target of brain damage and age-related diseases such as depression, Alzheimer’s Disease (AD), and Parkinson’s [1,2]. Gender has a different effect on the prevalence, manifestations, and effectiveness of treatment methods for these kinds of diseases. AD is a progressive neurodegenerative disorder of the brain that involves impairment of hippocampal integration and cognition [3]. The Cornu Ammonis (CA1) region of the hippocampus and the entorhinal cortex, which send afferent inputs to the dentate gyrus (DG), are the first brain regions to be affected by AD.
The DG of the hippocampus maintains the ability to generate new neurons in adulthood. On the other hand, adult hippocampal neurogenesis (AHN: Adult Hippocampal Neurogenesis) is closely related to learning and memory [4]. These new neurons can modulate the function of the hippocampus, which is a different form of learning and memory [5]. Due to the important role of these two hippocampal regions in cognition, memory, and learning, investigating the neural plasticity and the density of nerve cells and dendritic spines that play a role in synapse formation and neuron function seems necessary. Indeed, impaired neurogenesis may have important implications for the cognitive deficits seen in AD. Gender differences in the morphology and function of the hippocampus are evident and also affect how the hippocampus is involved in this disease. In addition to neurogenesis, the hippocampus changes under the influence of sex, sex hormones, dendritic architecture, synapse density, synaptic plasticity, and neurogenesis in adults [5]. The results of studies have shown that the risk of AD in women is higher than that of men, even taking into account the longer life span [5,6].
It seems that by replacing steroid hormones, especially estradiol, degenerative changes can be prevented to some extent or treated if they arise. Estrogen, as one of the sex hormones with its effect on the growth, differentiation, maturation, and functioning of the central nervous system, plays an important role in the functioning of this system [7].
In previous studies, we investigated different aspects of the sex hormone’s effect on neurogenesis in the hippocampus and subventricular zone. On the other hand, the effect of 17-beta estradiol treatment on the hippocampus in the ovariectomy model was mentioned. We also discussed the role of sex hormones on hippocampal astrocytes and the expression of neurotrophic factors. Therefore, our aim in the present study was to investigate the role of estrogen hormone on the morphology and plasticity of the hippocampus by the Golgi staining technique to show the density of nerve cells and dendritic branches. It is assumed that the morphological changes in the neurons of the DG and CA1 regions of the hippocampus are related to memory and learning. Cognitive disorders such as Alzheimer’s disease are thought to be related to AHN, and regulation of AHN can be used as a suitable therapeutic strategy to delay the progression of these disorders.
Animals and groups
In this experimental study, female NMRI laboratory mice 6-8 weeks were used. Mice were obtained from the Razi Vaccine and Serum Research Institute. They were housed in the animal lab of Damghan University under suitable conditions of 12 hours light/12 hours darkness, 20-24°C temperature, and with sufficient water and food. Mice were divided into 4 groups based on their vaginal smears: proestrus (n=7), estrus (n=7), metestrus (n=7), diestrus (n=7), and the 5th group was the ovariectomy group (n=7). To prepare a vaginal smear, after transferring the vaginal secretions on the slide, the samples were dried at room temperature, fixed with methanol, and stained with 2% methylene blue.
Bilateral ovariectomy
After determining the estrus cycle, mice in the diestrus phase were selected, because in this phase the cell composition is similar to the ovariectomized model and the level of ovarian hormones is at its lowest. Then based on their weight, a combination of ketamine-xylazine (K, 100 mg/kg; X, 10 mg/kg) was administered Intraperitoneally (IP). After shaving the animal’s hair with a scalpel blade, a small incision was made on the skin of the middle region of the back, and it was separated from its underlying tissues. The peritoneum was cut, and after visualizing the ovaries, they were completely separated with sterile scissors. The suture site was smeared with penicillin antibiotic and the weight of the mouse was measured before and after surgery [8].
Golgi staining
After sacrificing the mice, their brains were extracted and placed in the prepared Golgi solution in dark for one week. Afterward, the brains were cut with a vibratome with a diameter of 50 micrometers. Then, the slices of the hippocampus were placed in a 10% ammonia solution. Several sections of each sample were selected and prepared for histological examination. The cells in each section were observed using a light microscope.
Cresyl violet staining
After anesthetization, the animals were perfused by intracardiac injection of phosphate buffer solution followed by 4% paraformaldehyde solution. Then, the brains were removed, fixed, processed, and embedded in molten paraffin. Using a rotary microtome, coronal sections of 5 micrometers were prepared based on the Paxinus atlas in the DG region. The Cresyl Viola method was used to stain Nissl bodies of nerve cells. Afterward, the granular cells were counted in serial sections at each 10th section.
Estrogen hormone measurement by ELISA
To measure hormones, blood was collected from rat hearts in different stages of the estrus cycle. The blood serum was separated by centrifugation and kept at -70°C. Estrogen hormone was measured by ELISA according to the Dia Metra Company’s instructions [9].
Statistical analysis
Using image J software, neuron density, extra branching, and staining were evaluated. The obtained data were analyzed using a one-way analysis of variance and Tukey’s test. The results were presented as mean ± standard error and p< 0.05 was considered as a significant indicator.
Neuron density, nerve branches, and dendritic spines in the CA1 and DG regions by Golgi staining
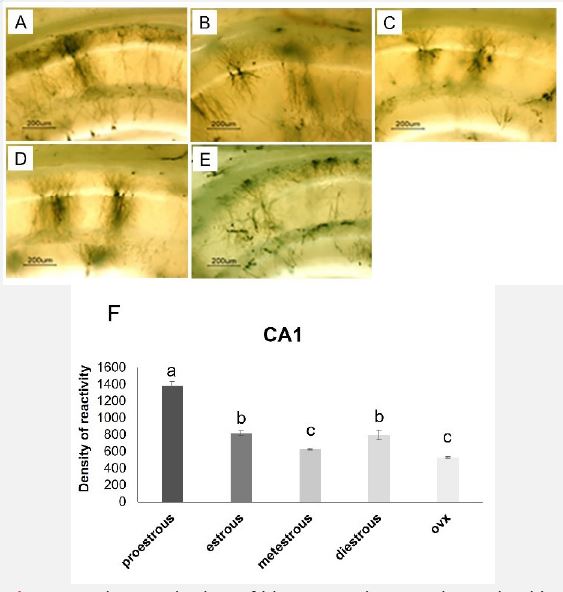
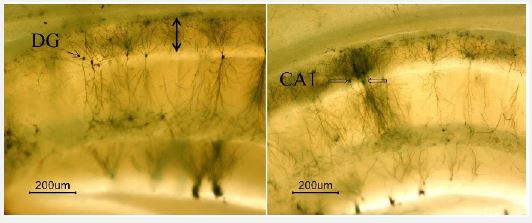
In previous studies, vaginal smear results have been reported to determine the estrous cycle [10]. As can be seen in Figure 1A-1J, the comparison of the neurons of the Dentate Gyrus (DG) in the proestrus phase, compared to other stages of the estrous cycle and the ovariectomy group, in terms of cell density, staining, and nerve branches is different. The amount of immunoreactivity, cell density, and branching of neurons in the CA1 region is also higher in the proestrus phase compared to the other groups and the ovariectomy group (Figure 2). In the proestrus stage, the branching of neurons can be seen with a microscope. The color density of neurons, dendritic branches, and the clarity of neural appendages in the images indicate the maturity of most nerve cells (Figure 3). Turkey’s supplementary test also shows a significant difference in neuronal density, extra branching, and staining of neuron in the microscopic sections of the proestrus group compared to other groups (Figure 1K and 2K) (p< 0.05).
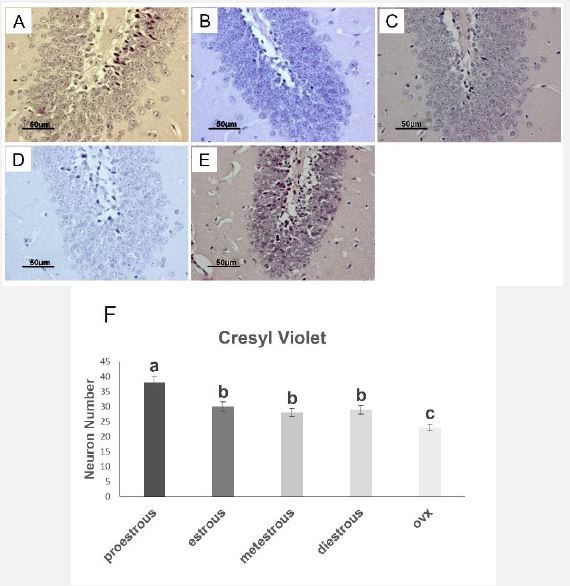
Hippocampal DG region by cresyl violet staining
In the proestrus group, the density of neurons of the granular layer of the hippocampus showed a significant increase compared to other groups. No significant difference was observed in the estrus, metestrus, and diestrus groups. In the ovariectomy group, neuronal density revealed a substantial decrease compared to other stages (Figure 4A-F).
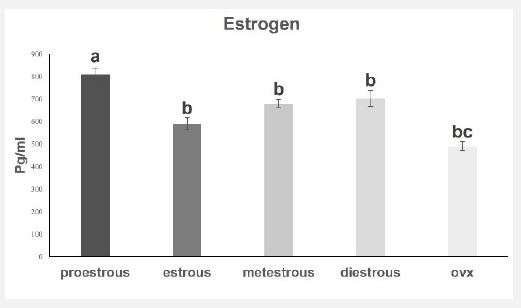
Estrogen concentration
As can be seen in Figure 5, the examination of the level of estrogen hormone in the blood plasma showed that the concentration of estrogen in the proestrus group showed a significant increase compared to other groups (p<0.05). Turkey’s supplementary test showed that estrogen concentration in estrus, metestrus, and diestrus groups did not show any significant difference from each other. In the orchiectomy group, the concentration of estrogen is significantly reduced compared to other groups (p<0.05).
The results of this research showed that in the proestrus phase of the estrous cycle when the plasma concentration of estrogen hormone was the highest in the blood plasma, the highest reactivity of dendritic spines and growths and the density of nerve cells in the DG and CA1 regions were observed in the hippocampus.
The hippocampus plays an important role in learning and responding to stress and therefore plays a vital role in complex and innate behaviors. Aspects of learning and response to stress in men and women are different throughout life, and disorders related to hippocampus dysfunction, such as depression, anxiety, or schizophrenia, differ between sexes in terms of prevalence or manifestation [11,12]. While evidence suggests that many of these secondary differences are related to the developmental organizational effects of gonadal steroids, the mechanisms of which are unknown [13]. Previous studies showed that gonadal steroids cause sex differences in cell production in the hippocampus of newborn rats [14-16]. Extensive research has been done on the effect of sex steroid hormones on the brain. The results of these investigations show that sex hormones exert a wide range of neuroprotection [17], including anti-apoptotic and antioxidant properties, as well as increasing neurogenesis and synaptic plasticity in the brain [12]. Steroids are factors that determine the proliferation [18], differentiation, and organization of the central and peripheral nervous systems. Steroids can distinguish the structure and function of the brain during life [19]. Also, estrogens affect the plasticity process in multiple brain regions through the genomic and non-genomic Estrogen Receptor alpha (ERα) signaling pathway [20].
Neuroplasticity in the hippocampus, in addition to the controlled production of new neurons and their maturation, which is called neurogenesis, also includes the modification of synapses and dendrites [21]. In this research, it was reported an increase in dendritic branches as well as synaptic density in DG and CA1 as a sign of neuroplasticity and neurogenesis. Barrera and Markham showed that in men, the number of dendritic branches in the granule neurons of the dentate gyrus and CA1 pyramidal neurons is higher than in women [22,23]. Since two factors of cognitive decline and hippocampal morphology are affected by sex hormones, Markham using the Golgi technique in young adult rats showed that the dendritic branches of CA1 neurons are different [23]. Therefore, in the present study, the Golgi staining method, which is a well-known method to show the branching of nerve cells and the density of dendritic or synaptic spines (as an important morphological indicator to show synaptic activity) and nerve maturity, was used [24]. In another study, it was observed that using YL-0919 quickly improved anxiety and depression-like behavior in mice in one week. They also compensated for the decrease in hippocampal dendritic formations and dendritic spines caused by estrogen withdrawal [25]. In this study, we observed an increase in pigmentation and dendritic and synaptic branches in the estrogen treatment group compared to the ovariectomy group (Figures 1 and 2). In other studies, estrogens were shown to modulate multiple synaptic plasticity processes in the CA1 region of the hippocampus. Estrogens in vitro and in vivo increase dendritic spine density and synaptic proteins in CA1. Also, estrogens regulate the expression of TrkB neurotrophin receptor in dendritic spines and terminals in CA1 and DG, and TrkB is important for long-term potentiation and modification of dendritic spine structure [20].
The estrus cycle in mice occurs for four to five days and includes four stages proestrus, estrus, metestrus, and diestrus. Endocrine events, ovarian changes, and the amount of vaginal mucus secretion change during the different stages of the estrous cycle [26,27]. At the time of ovulation (the end of proestrus and the beginning of estrus), the plasma concentration of estrogen reaches its peak. The plasma concentration of estrogen is low in metestrus and diestrus, and the concentration of estrogen gradually increases at the end of diestrus [28-30]. In rodents and virological conditions, the number of gonadal hormones changes in a short time. The high level of endogenous and exogenous estradiol in female rats is related to the increase in the cell proliferation rate of the dentate gyrus. The proestrus stage of rats has a higher cell proliferation rate than the stages of estrus and diestrus [31,32]. In another study, the density and number of proliferated cells in the dentate gyrus region of the rat hippocampus in the proestrus stage were reported to be more than the other stages of the estrous cycle. Also, in another study, the morphology of astrocytes and the response to the GFAP marker were different in all stages of estrus. In addition, at different stages of the estrous cycle, differences in the morphology of astrocytes and the amount of GFAP staining were observed [33,34]. Galea et al also reported differences in neurogenesis during the estrous cycle [35]. In rodents, due to the short estrous cycle, gonadal hormones change significantly in a short time. For example, during diestrus, the amount of circulating estradiol is low, but it is high in proestrus. The difference in the amount of estrogen during the estrous cycle may change the morphology and number of brain cells [36]. In a study, researchers found that after injecting Bromodeoxyuridine-BrdU into rats in the proestrus stage (highest estrogen levels), the rate of cell proliferation and survival in the dentate gyrus region of the hippocampus showed a greater increase than in other estrus stages. In this regard, with the decrease in circulating estrogen levels and the number of estrogen receptors, in the short term, the proliferation of dentate gyrus cells in ovariectomized female rats has also decreased [37]. This finding is consistent with our previous studies and other studies that showed that cell proliferation was higher in the hippocampal dentate gyrus of the proestrus stage and ovariectomized rats treated with 17-beta-estradiol [33,38-40]. 17β-estradiol levels change during the menstrual cycle in mammals (or the estrous cycle in rodents) and throughout life [41,42]. Significant changes in estrogen levels throughout life can affect hippocampal neuroplasticity, cognition, and brain volume. These changes are related to estrogen exposure during life, increasing age, and the type and duration of Hormone Therapy (HT) [43-45].
During menopause, there is a significant decrease in the levels of 17β-estradiol and estrone [46]. Studies have shown that the early onset of menopause in women (either naturally or surgically induced) is associated with an increased risk of AD. Menopause caused by surgery compared to natural menopause is associated with a faster decline in verbal and episodic memory performance [47,48]. Estrogen, as one of the sex hormones, plays an important role in the development and function of the Central Nervous System (CNS), including the regulation of mood, emotions, mental states, learning, memory, cognition, and neurodevelopmental processes. There is considerable evidence that estrogen can play a protective role by modulating neuronal growth, neuronal excitability, dendritogenesis, and synaptic plasticity [49]. Therefore, this study, along with other studies, can emphasize the neuroprotective effect of estrogen to delay the onset and spread of cognitive disorders as an alternative therapy (ERT).
The results show that sex hormones, especially estrogen, are effective on neuronal density and dendritic branches in the DG and CA1 regions of the hippocampus, and this effect can subsequently affect cognition, memory, and learning.
Funding sources: The authors did not receive any funding for this research study.
Conflict of interest statement: All authors have no any potential declarations of interest.
Acknowledgments: The authors thank the Faculty of Biological Sciences, Damghan University (Damghan, Iran) for supporting perform this research article.
Statement of ethics: This study protocol was reviewed and approved by Animal Ethics Committee of Department of Cellular and Molecular Biology, School of Biology, Damghan University, Damghan, Iran, approval number 178-9/4/1393.
