1Department of Tech. Biosciences, Digianalix, South Samaj Street, Tharpakhna, Ranchi-834001, Jharkhand, India.
2Department of Biotechnology, Marwari College Ranchi-834001, Jharkhand, India.
3Department of Biotechnology, St. Xavier College, Ranchi-834001, Jharkhand, India.
Mukesh Nitin
Tel: +917859045686;
Email: digianalix@gmail.com
Received : Oct 19, 2023 Accepted : Nov 23, 2023 Published : Nov 30, 2023 Archived : www.meddiscoveries.org
Background: Typhoid is a life threatening disorder caused by Salmonella enterica typhi has been a crucial topic of study in developing and underdeveloped countries especially India, some Asian, African and Middle Eastern countries which are reported to be affected with this bacteria. Around 14 million cases were observed in recent years. As, S. typhi bacteria is dispersing its roots through contaminated water and foods across the globe. With the advances in computational biology and next generation sequencing we identified different genomics data of variable typhoid cases across the globe, through the meta-analysis approach. We had screened the potential top 20 hub genes showing their significant expression (p-value≤0.05) in multiple genomic typhoid disorder cases. This identification resulted in uncovering vital metabolic pathways that can be further used to highlight specific target receptors like AKT1 and CD4, playing essential role in variable typhoid conditions.
Methods: We used drug design and development approaches to identify essential natural compounds, library preparation based on extensive bibliographic literature survey and ADMET properties analysis using CHIMERA, Autodock, Osiris and DruLito for finding potential candidate drug. Further, molecular dynamic simulation were carried out using iMOD server.
Results: Molecular docking was carried out between the potential target receptors and the ligands resulted in the identification of receptor AKT1 and Marmesinin having docked score (ΔG-8.0) as test and ceftriaxone having docked score (ΔG-6.6) as control. CD4 and formyl steroids having docked score (ΔG-6.3) as test and ciprofloxacin having docked score (ΔG-5.3) as control followed by MD simulation.
Conclusion: Marmesinin, a natural bioactive compound manifests the best docking result with both the target receptors. Furthermore, our study reflected that in future marmesinin can be used as a potential drug target for typhoid treatment under multiple typhoid disorders across the globe.
Keywords: Typhoid; Next generation sequencing; Hub genes; Receptor; Docking; Marmesinin.
Typhoid fever cause majorly by Salmonella enterica typhi and has posed its roots resulting in life threatening death across the globe. Recently, it was reported that approx. 14.3 million cases occurred due to typhoid and para typhoid in countries like India, Pakistan, Bangladesh, Vietnam and South Asia countries, which includes various Southern pacific, African and some countries of middle east [1]. In this growing world of processed food, humans came across numerous hazardous infections. Amongst all these infections, Salmonella is known as major threat because of consumption of pork or pork products. According to the European Union cases of food borne outbreaks are frequently noticed for the Salmonella resulted in the alternation of mRNA and its protein expression [2,3].
As S.typhi is an obligate, gram-negative pathogen and is majorly reported for inducing infection in humans as the only host. Despite of consuming processed food, ingestion of contaminated food or water is also an aspect through which humans acquired infection. This type of compromised food chain dependent microorganism can be seen in the areas with no or least clean water, non-hygienic life conditions and sanitation system [4,5]. Elimination or restricting the diseased condition, improvement in quality of drinking water and sanitation conditions are essentially required. But unfortunately, these interventions are not promptly implemented in endemic areas of Africa and South Africa [6].
There are 2 methods through which transmission of this disease takes place short cycle- in this cycle is mainly related to food handlers and got transmitted within the house hold [7]. Long cycle- it occurs through contamination and contact with external contaminants. The behavioral role of this pathogen were reported to behave as a feral matter for one host and are consumed by another. There is also survival of S. typhi outside the human body through amoebas. Through survey, reported that in Kathmandu, Nepal contamination of drinking water through stope taps cause major transmission this pathogen [8,9].
Currently, pharmaceutical approaches provided two licensed vaccines for typhoid but they are effective up to moderate level i.e., 50-55%. These medicines are not profusely practiced in endemic regions. Also, the reports on bacterial behavior against available typhoid vaccine is still in its limited effect thus representing a serious threat to human health. So, it is the vital need to develop or improve of current medicinal drugs to combat against typhoid. For the development of potential and effective vaccine, better understanding of protein or target antigens are required. This would aid individual’s immune system to maintain and boost the expression of target antigens. Due to this action, it could be helpful to incorporate to form new multivalent or conjugate subunit vaccines [10,11]. The synthetic drug used for the cure of this endemic disease comes with numerous side effects which includes constipation, excitement and coma. They also cause effects in immune system like hypersensitivity and drug interactions. So, the phytochemical from natural source would be better alternative to cure this disorder for long term [12,13].
With the advancement in next generation sequencing and bioinformatics applications we can develop the potential natural drug component which may play a crucial role in targeting the cure of typhoid disease. Moreover, the study of gene expression behavior using NGS will open a new dimension in the exploration of potential gene contribution in human body to counter the bacterial infection targeting typhoid. In our research study we proposed a comprehensive pipeline and metaanalysis approach that leverages the heterogeneity on widely explored experimental data set in order to identify the robust, reproducible disease potential gene signatures. We also tried the meta-analysis of DNA microarray expression datasets on various typhoid cases to decipher the patterns of genes which are commonly and collectively responsible for these disorders. This research article will add value to scientific communities for the preparation of next level road map resulting in the identification of potential hub-genes showing conservancy in various typhoid disorders and can be further used in drug discovery studies for designing of potential drug to cure majorly chronic diseases like typhoid through docking studies.
Sample collection
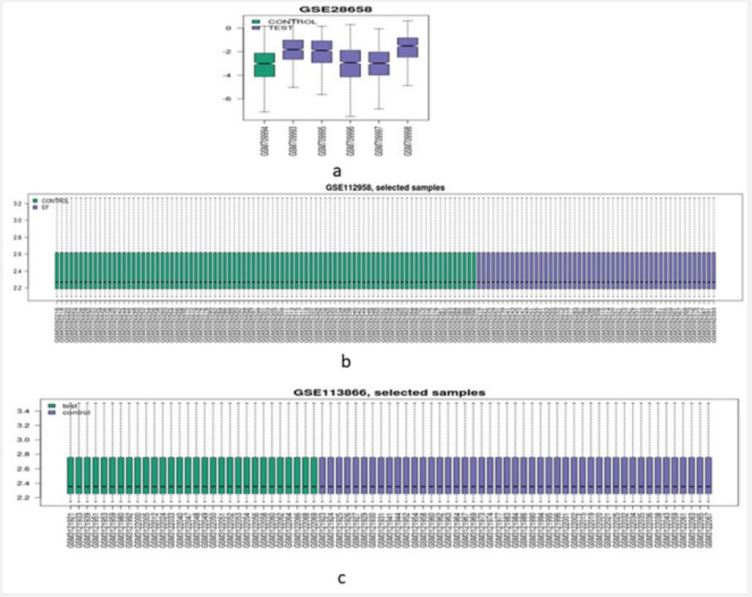
During our study various genomics based DNA microarray datasets on typhoid were collected from public available NCBI-Gene Expression Omnibus (GEO) database, where different cases on typhoid were observed and screened for our research. In our analysis we considered three different datasets of variable typhoid study represented as case 1 (GSE28658) highlighting expression analysis of data from infected patients of typhoid and infected patients from other bacteremic from Nigerian children containing 6 samples, case 2 (GSE112958) show the expression of whole blood of healthy adult volunteers experimentally challenged with S. Typhi containing 178 samples and case 3 (GSE113866) expression of febrile culture confirmed enteric fever cases containing 149 samples. The selected datasets were categorized on the basis of two parameters TEST and CONTROL from the uninfected and healthy samples of typhoid.
Meta-analysis and gene ontology
The datasets were re-analyzed using GEO analyzer NCBI server [14]. Further, the analysis resulted in the generation of box plot, volcano plot and MA plot along with gene expression table using Benjamin-Hedge Hoch test, Limma package-R software [15]. The selected datasets were meta-analyzed and sorted on the basis of P<0.05 using shell script. The different expressed genes in datasets were screened and they were further analyzed in Venn Ghent server [16] to explore the conserved genes present in them. The common obtained genes from conservancy analysis were further imported to STRING (search tool for the retrieval of interacting genes) [17], to study the protein -protein interaction network. The screening of resultant proteins were further exported into Cytoscape for visualization. The genes were screened by using String app, Cytohubba [18] and y- files algorithm to identify the hub genes network. The screened hub genes which played crucial role in futuristic target prediction were subjected to gene ontology using Gene ontology server to highlight metabolic pathways [19].
Homology modelling and receptor preparation
The two hub genes AKT1 and CD4 were screened for homology modelling. The sequences were imported from UNIPROT [20] and physiochemical properties were evaluated using the ProtParam tool [21], secondary structural prediction in SOPMA [22], transmembrane properties using SOSUI server [23] and Ramachandran plot using Procheck [24]. AKT1 and CD4 were further used for receptor preparation with UCSF Chimera [25]. The already present ions and ligands from the modelled protein structures were screened and removed. The hydrogen and charge were added, surface binding and dock preparation were accomplished in final PDB format for further analysis.
Screening of bio-active compounds and ligand preparation
An in-depth literature study were carried out to identify bio active compounds present in plants and herbs, which were observed crucial in the treatment of various endemic disease like typhoid. Those compounds were then selected and downloaded from PUBCHEM [26] leading into library preparation ADMET properties for the candidate ligand selection were carried out using DruLito [27] and OSIRIS [28] to check the toxicity such as irritant, mutagenic, tumorigenic and reproductive effect. The compounds which passed toxicity risk were discarded and the remaining ADMET checked passed ligands were further screened out as our potential target ligand for docking studies. Avogadro software [29] was used by the potential ligands for the force filed and geometry optimization and were further subjected to USCF chimera for addition of ion and charge to pass final ligand preparation analysis.
Molecular docking studies
The receptors AKT1 and CD4 along with final ligands were further subjected for the docking studies in CHIMERA-Auto dock Vina [25]. The docking studies revealed that the docked complexes show a high negative binding energy score ΔG which indicates the principle of minimum energy and high stability leading to best interaction between proteins and the ligands. The obtained ligand-receptor docked complex were further visualized in Biovia Discovery studio [30] to interpret docking analysis between the bonds of ligands and receptors.
Genomics and Meta-analysis of samples
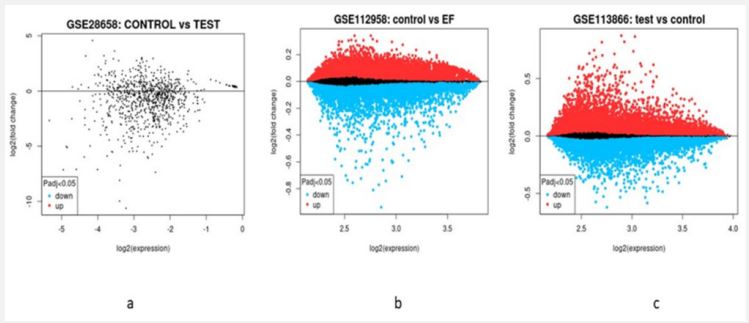
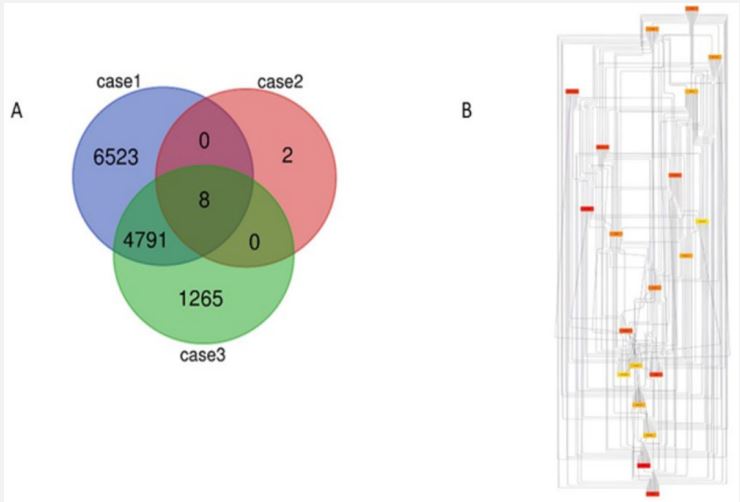
Typhoid poses a serious which is affecting people worldwide. During our study we used three variable typhoid cases which were downloaded from NCBI dataset and were further analyzed using genomics approaches to identify multiple genes expression analysis. All the samples were subjected to read normalization prior to genomic analysis as shown in (Figure 1). The significant differential expression analysis having (p-value≤0.05) for test and controlled samples of typhoid cases resulted in the identification of total 907, 47231, 47232 genes in Case 1 (GSE28658), Case 2 (GSE112958), Case 3 (GSE113866) represented as volcano and MA plot (Figure 2 and 3). The qualified set of significant genes across the three cases were imported for gene conservancy analysis in order to highlight the most commonly expressed set of genes present in three diversified cases of typhoid disease. It resulted in the identification of total 4799 conserved genes from all the three cases, and were parsed for the identification of potential hub genes through protein-protein interaction network study with STRING algorithm. From the pool of conserved genes, we identified 20 most expressed hub genes like AKT1, CD4, IL1B, STAT3, ALB, etc. as shown in (Figure 4).
Pathway analysis and target receptor preparation
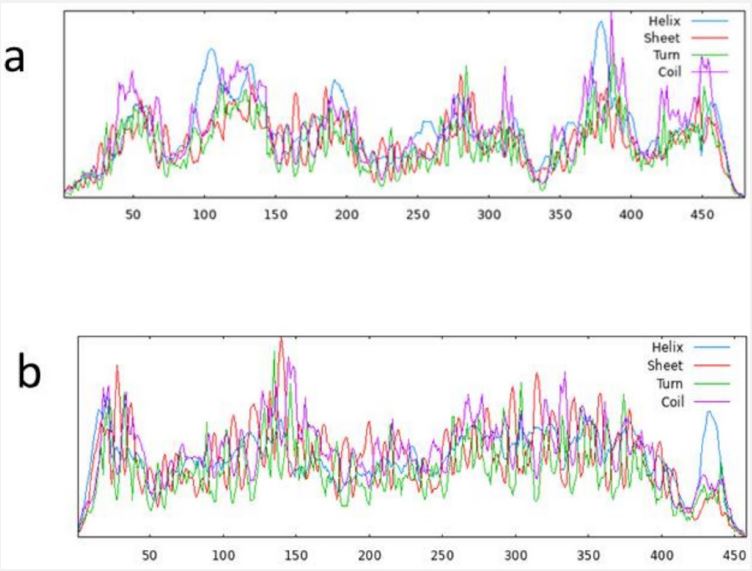
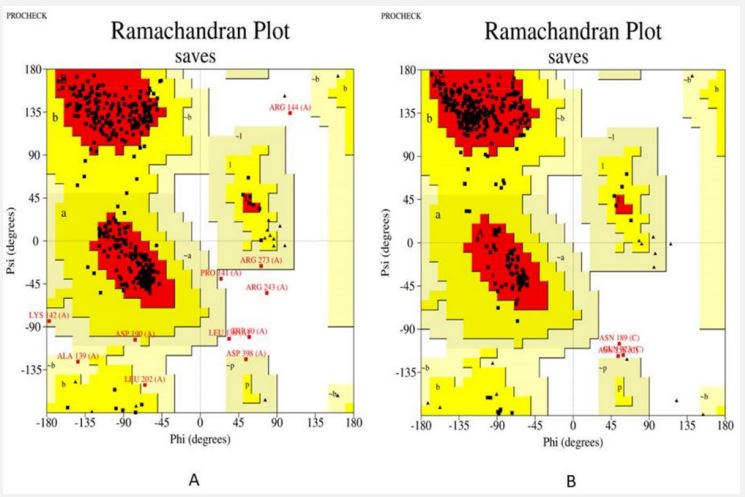
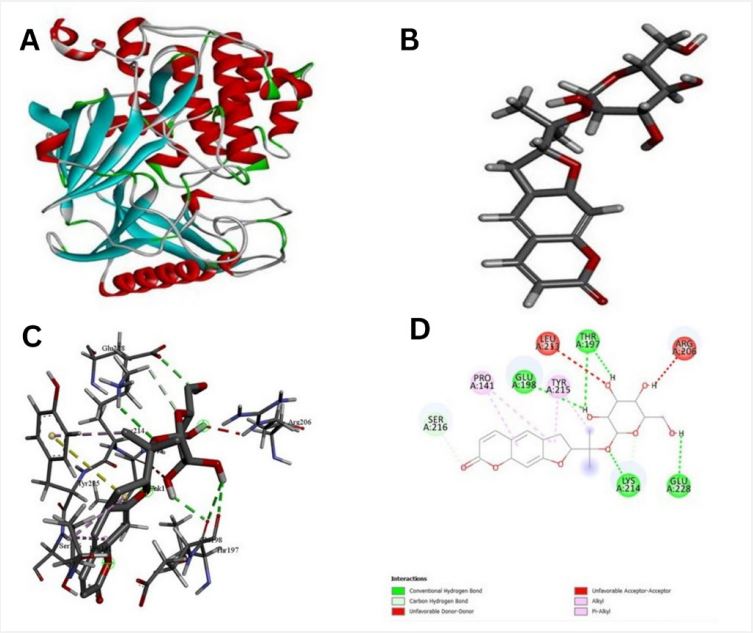
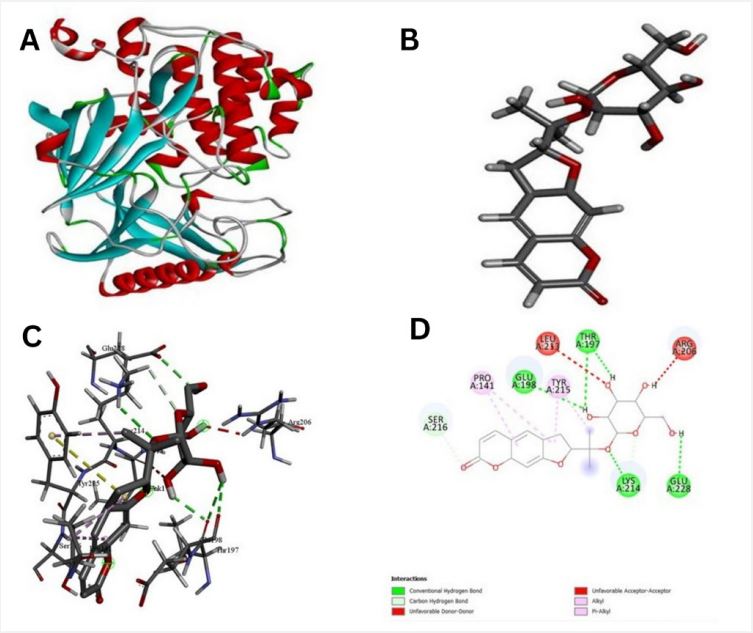
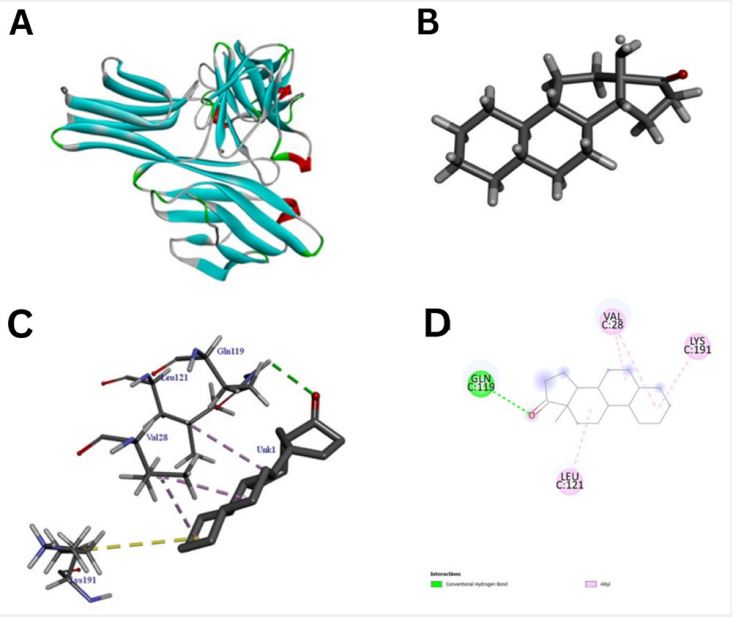
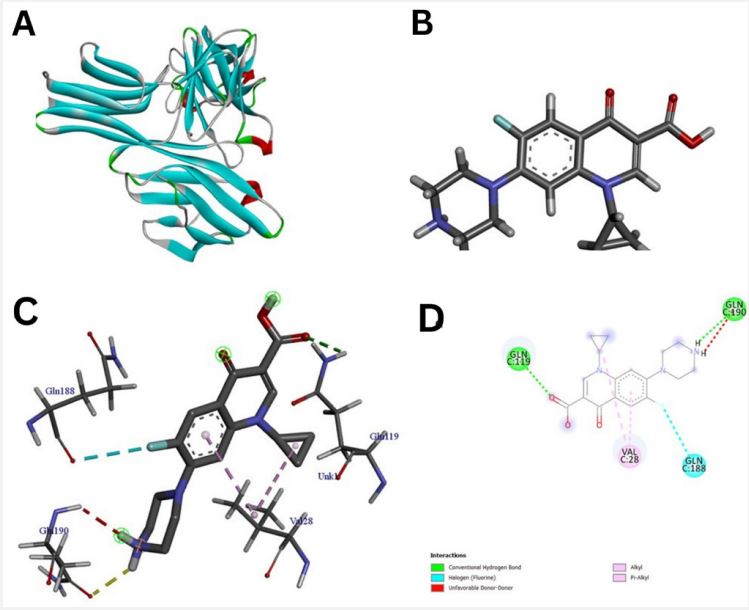
The pathway of resultant hub-genes was studied based on multiple literatures as references which helped us to conclude that the genes are useful in pathways related to typhoid, cancer and many other cellular processes. Among 20 hub genes, two genes AKT1 and CD4 were selected for the further study due to their close relatedness with the typhoid and defined metabolic pathway (Figure 5). Further, homology modeling was carried out followed by physio-chemical structural analysis of AKT1 and CD4 were carried out and showed instability index of 35.47 and 41.78, hydrophobicity (GRAVY) of -0.575 and -0.257 respectively. The genes were further analyzed to check secondary structure analysis of a protein and detection of helix which resulted in identification of length of AKT1 as 480 and similarity threshold value 8 and length of CD4 as 458 and threshold value as 8 as shown in (Figures 6 and 7; Table 1 and 2). Ramachandran plot of AKT1 and CD4 were plotted showing favored regions 90.3%, 91.0% and additional allowed regions 8.1%, 7.1% as shown in (Figure 8 and Table 3). The potential genes AKT1 and CD4 were imported for receptor preparation by removing additional chains, ions and solvents. Using SPDV the receptors were carried out for energy minimization followed by addition of hydrogen and charge in GAFF force field for final target receptor preparation.
Library preparation of bio-active compounds
A total of 38 metabolites were selected from herbal plants to study their medicinal properties to cure typhoid through several research and review papers further the metabolites were passed to check their ADMET properties wherein metabolites were filtered on parameters like cLog P, solubility, molecular weight, TPSA, drug likeliness and drug score. Out of total 16 ligands passed the screening process like Marmesinin and steroids having 0.02, -2.8, 408, 134.9, -7.1, 0.41 and 4.12, -4.55, 260, 17.07, -4.67, 0.34 respectively showing no toxicity such as irritant, mutagenic, tumorigenic and reproductive effect. These 16 metabolites were- Marmesinin, Beta–sitosterol, Chebulic acid, Gallotannin, n-hexacosane, S-Allylcysteine, Glycylrrhizin, Saponin, Lycopene, Alliin, Diallyl Disulphide, Cholic acid, Methyl inositol, Alpha-Glycyrrhizin, Steroids and Formyl steroid as shown in (Supplementary Table 1 and 2).
Docking studies and MD simulation
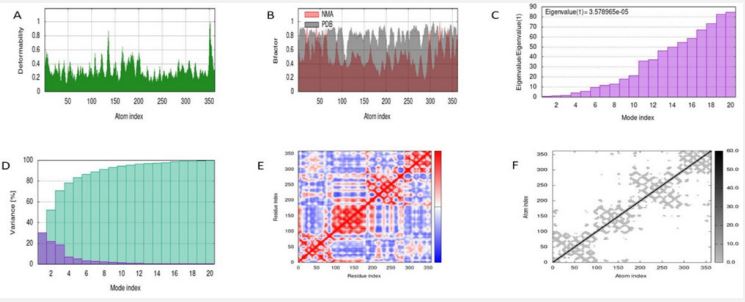
The resultant 16 ligands with 2 potential target receptors were selected for molecular docking studies (Supplementary Table 3). On the basis of values for pocket id for AKT1 as X=10.256, Y=-14.157, Z=5.086 and respectively for CD4 as X=-16.559, Y=-35.147 and Z=31.379 with center size of 20. Molecular docking of Marmesinin (73191) with AKT1 interacted with the binding energy of (ΔG) -8.0 and docking of steroids with CD4 as (∆G) -6.1 whereas docking score of AKT1 and CD4 were carried out against standard FDA approved drugs showing AKT1 with cefixime (ΔG) -7.1 and CD4 with ciprofloxacin (ΔG) -5.3 as shown in (Figures 9-12 and Table 4). The docked complexes were then visualized to get a brief about bonds and interactions among them showing AKT1-Marmesinin H-bond(2.74A0 , 2.87 A0 , 2.63A0 , 2.23A0 , 2.31A0 , 2.75A0 , 2.60A0 ), Hydrophobic bond (4.69A0 , 5.21A0 , 4.84A0 , 5.36A0 ) Intrinsic interactions bond (2.26A0 ) and AKT1-Steroids Showing H–bond (2.80A0 ) and Hydrophobic bond (4.87A0 , 4.24A0 , 4.27A0 , 5.38A0 ) as shown in (Table 5, Supplementary Table 4, Figures 1-12). Molecular dynamics simulation was then carried out for the best docked complex against both the receptors AKT1 and CD4 with ligand Marmesinin showing the Eigen value of1.451971e-04, 3.578965e-05, Deformability hinges 5(0.6,0.6,1,0.8,0.7), 4(0.5,0.8,0.6,1) respectively. We also observed that the favorable region along with steepness is high around the base line for both docked complexes. Further the dynamics simulation was visualized based on b-factor, variance and residue plots as shown in (Figures 13 and 14).

| SOPMA | AKT1 | CD4 |
|---|---|---|
| Alpha helix | 36.04% | 19.65% |
| 310 helix | 0.00% | 0.00% |
| Pi helix | 0.00% | 0.00% |
| Beta bridge | 0.00% | 0.00% |
| Extended strand | 17.08% | 31.00% |
| Beta turn | 7.29% | 7.21% |
| Bend region | 0.00% | 0.00% |
| Random coil | 39.58% | 42.14% |
| Ambiguous states | 0.00% | 0.00% |
| Other states | 0.00% | 0.00% |
| No. | Receptor | Region | Transmembrane sequence | Type |
|---|---|---|---|---|
| 1 | AKT1 | 103-124 | CCEEEECCCCCCCCCCCCCCCC | Primary |
| 2 | 907-929 | CCCCCCCCCCCCCHHCCCCCCCC | Primary | |
| 3 | CD4 | 4-25 | GVPFRHLLLVLQLALLPAATQG | Secondary |
| 4 | 545-567 | CCCCCCCCCCCCCCCEHLTLPQA | Primary | |
| 5 | 746-768 | PMALIVLGGVAGLLLFIGLGIFF | Primary |
| Ramachandran plot analysis | AKT1 | CD4 |
|---|---|---|
| Residues in most favoured regions [A,B,L] | 90.3% | 91.0% |
| Residues in additional allowed regions [a,b,1,p] | 7.1% | 8.1% |
| Residues in generously allowed regions [~a,~b,~1,~p] | 1.5% | 0.0% |
| Residues in disallowed regions | 1.0% | 0.9% |
| Sr.no | Receptor | Compound | Score | RMSD I.b. | RMSD u.b. |
|---|---|---|---|---|---|
| 1 | AKT1 | TEST | |||
| Marmesinin(73191) | -8.0 | 0.0 | 0.0 | ||
| CONTROL | |||||
| Cefixime(5362065) | -7.1 | 0.0 | 0.0 | ||
| 2 | CD4 | TEST | |||
| Steroids(4226429) | -6.1 | 0.0 | 0.0 | ||
| CONTROL | |||||
| Ciprofloxacin(2764) | -5.3 | 0.0 | 0.0 |
| Receptor-LigandInteraction | Vina score/Binding affinity (kcal/mol) | Ligands | Distance(A0) | Receptor | Category | Types of Interaction |
|---|---|---|---|---|---|---|
| AKT1 – Marme- sinin (Test) | -8.0 | UNL1:H | 2.74 | A: GLU228: OE2 | H-bond | Conventional bond interaction |
| UNL1:O | 2.87 | A: LYS214: HE2 | H-bond | Carbon – H bond interaction | ||
| UNL1:O | 2.63 | A: LYS214: HZ1 | H-bond | Conventional bond interaction | ||
| UNL1:C | 4.69 | A: TYR215 | Hydrophobic | Pi-Alkyl interaction | ||
| UNL1 | 5.21 | A: TYR215 | Hydrophobic | Pi-Alkyl interaction | ||
| UNL1 | 4.84 | A: PRO141 | Hydrophobic | Alkyl interaction | ||
| UNL1 | 5.36 | A: PRO141 | Hydrophobic | Alkyl interaction | ||
| UNL1:O | 2.23 | A: SER216: HB1 | H-bond | Carbon – H bond interaction | ||
| UNL1:H | 2.31 | A: THR197: O | H-bond | Conventional bond interaction | ||
| UNL1:H | 2.75 | A: THR197: O | H-bond | Conventional bond interaction | ||
| UN1:H | 2.60 | A: GLU198: O | H-bond | Conventional bond interaction | ||
| UNL1:H | 2.26 | A: ARG206: HE | Intrinsic interactions | Unfavourable donor-donor interaction | ||
| AKT1- Cefixime (Control) |
-7.1 | UNL1:N | 4.02 | A: LYS214: NZ | Intrinsic | interaction Unfavourable positive-positive interaction |
| UNL1:S | 5.56 | A: LYS214: NZ | Intrinsic interaction | Unfavourable positive-positive interaction | ||
| UNL1:N | 2.37 | A: LYS214: HZ1 | H-bond | Conventional bond interaction | ||
| UNL1:H | 2.96 | A: LYS214: O | H-bond | Conventional bond | ||
| CD4- Steroids (Test) | -6.1 | UNL1:O | 2.80 | C: GALN119: HE22 | H-bond | Conventional hydrogen bond interaction |
| UNL1 | 4.87 | C: LEU121 | Hydrophobic | Alkyl interaction | ||
| UNL1 | 4.24 | C: VAL28 | Hydrophobic | Alkyl interaction | ||
| UNL1 | 4.27 | C: VAL28 | Hydrophobic | Alkyl interaction | ||
| UNL1 | 5.38 | C: LYS191 | Hydrophobic | Alkyl interaction | ||
| CD4 – Ciprofloxacin (Control) | -5.3 | UNL1:F | 3.59 | C: GLN188: O | Non-covalent interaction | Halogen(fluorine) Interaction |
| UNL1:O | 2.74 | C: GLN119: HE22 | H-bond | Conventional hydrogen interaction | ||
| UNL1:F | 3.59 | C: GLN188: O | Non-covalent interaction | Halogen(fluorine) Interaction | ||
| UNL1 | 4.57 | C: VAL28 | Hydrophobic | Pi – Alkyl interaction | ||
| UNL1 | 5.50 | C: VAL28 | Hydrophobic | Alkyl interaction | ||
| UNL1:HN2 | 2.48 | C: GLN190: HN | Intrinsic Interaction | Unfavourable donor-donor interaction | ||
| UNL1:H | 2.44 | C: GLN190: OE1 | H-bond | Conventional hydrogen bond interaction |
In recent years, typhoid disease has been a center of attraction and attention in different sites of medical research. In order to systematically analyze the mechanism of S. typhi bacteria in humans, experimental validation and in-depth studies were carried out. During our study using genomic approach we identified vital hub genes like ILIB, TLR4, SRC, MYC, JUN, AKT1, CD4 etc. after screening check of conservancy analysis across various typhoid cases. The similar study suggests that IL1B gets upregulated related to immune response during typhoid and play important role in salmonella infection [31]. TLR4 also reported in assisting in inhibition and recognition of S.typhi [32]. TNF a cytokine plays a major role in both pathogenesis and control of Salmonella infection. The genes SRC, MYC, JUN plays an important role in pathway of cancer and other diseases [33]. AKT1 and CD4 genes found in our analysis against typhoid diseases were considered as target receptors. These two hub genes AKT1 and CD4 were reported to play an important role in typhoid considering that AKT1 also modulates the function of numerous substrates, which regulates cell processes, such as metabolism, translation, proliferation, survival and angiogenesis [34]. The studies have shown the evidence of AKT1 as a promising candidate gene of the QTL region as the protein is known to be activated by Salmonella and to promote intracellular replication of the bacteria in mammalian cell. AKT1 gene may also play both positive and negative roles in control of salmonella infection [35], furthermore CD4 genes receptors helps in recognition and has a capacity to kill pathogen. CD4 also activates macrophages, B cells and other cells [36]. CD4 T cells are essential gene for protective immunity against oral Salmonella infection [37].
Further, we screened important bioactive compounds based on their medicinal properties to cure typhoid. Similar type of observations was reported using leaf extract of A. indica (neem) shows antibacterial activity against typhoid and para typhoid due to the presence of Steroids in them [38]. Higher and aromatic plants like mint used as one of the sources of traditional medicine from decades. This plant leaves are utilized in various herbal tea to add flavours and aroma. Methanol acts as important aromatic compound in mint plant showing its medicinal properties in curing gastrointestinal disorders, common cold and pain [39]. Also, this plant has a rich source of iron and magnesium that play a crucial role in human nutrition [40]. Thus, a total of 38 metabolites were selected from herbal plants in our study showing their medicinal properties to cure typhoid. Out of which 16 ligand compounds the best two like Marmesinin and Steroids passed the screening process of ADMET having properties of 0.02, -2.8, 408, 134.9, -7.1, 0.41 and 4.12, -4.55, 260, 17.07, -4.67, 0.34 for clogp, solubility, mol weight, TPSA, drug likeliness and drug score respectively. These ligands show no toxicity such as irritant, mutagenic, tumorigenic and reproductive effect. Docking studies were carried out for AKT1 and CD4 with respect to medicinal plant based natural compounds like Marmesinin and Steroids having docked score of (ΔG-8.0) and (ΔG-6.1) respectively whereas the FDA approved drugs like cefixime and ciprofloxacin were also used against receptors as control showing docked score of (ΔG-7.1) and (ΔG-5.3). With this docking score we found that medicinal compound Marmesinin resulted in best docked score comparatively against standard typhoid drug compound opening a new possibility for exploring more and effective plantbased compound studies which may play a crucial role in curing typhoid. Bioactive compounds like E1, E2, E3 and E4 (-10.3, -10.2, -9.9 and -10.6) also showed the best binding affinity compared to the standard antibiotic drug of typhoid ciprofloxacin (-6.6kcal) [41] similar to our finding where Marmesinin showed better docked score with respect to drug Ciprofloxacin. Similar type of compounds like Kaempferol and Silybin (group of flavonoids, polyphenols and alkaloids) medicinal plant source were also docked against various virus and bacterial diseases like SARS-CoV-2 protein which resulted in docked score of (ΔG8.3) and (ΔG-8.5) proving to be a potential drug candidate for covid [42]. Also, the use of medicinal plants like mint proved to be a potential drug candidate in typhoid. Similar to our research some other medicinal plant-based ligand compounds like Menthone, Beta pinene, Menthofuran and 1,8 cineole showed best docking results against L-lactate dehydrogenase. A chain, Retinal dehydrogenase, Lactate dehydrogenase and NADPH-dependent methylglyoxal reductase GRE [43]. Molecular dynamics simulation studies were carried out for our docked complex AKT1- Marmesinin and CD4-Marmesinin showing the eigen value of 1.451971e-04, 3.578965e-05 followed by deformability hinges 5(0.6,0.6,1,0.8,0.7), 4(0.5,0.8,0.6,1) and having favorable region along with high steepness based on b-factor and variance analysis. Similar type of MD studies was carried out for variable bacterial and lumpy disease which were also represented with favorable regions and high steepness [44,45]. The docked complex of our study signifies the possibility of finding potential natural drug compound that can be further curated in wet lab for eradicating typhoid based critical disorders.
Typhoid is still considered an important, trouble some issue for public health globally. Our, research analysis resulted in the identification of potential target genes AKT1 and CD4 based on its expression in variable typhoid disease. With molecular docking studies we identified natural bioactive compounds like Marmesinin and Steroids to be the most vital and dominant compound against AKT1 and CD4 receptors playing a crucial role in typhoid cases with respect to standard drug medicines. The present study also reflects that Marmesinin a bioactive compound obtained from Bael has a wide therapeutic property. It can be further validated in wet lab, assisting future researchers in proving it as a potential drug candidate to cure critical typhoid disorders across the globe.
Authors contribution
Daksh Kumar, Shweta Mishra, Annie Jessica Toppo, Md. Mahfooz Khan, Pramod Kumar, Aditi Kumari, Priyangulta Beck, Dipanjali Sharma, Kajal Kumari, Dinesh Kumar, contributed study, compiling, analysis of the manuscript; Daksh Kumar, Shweta Mishra, Md. Mahfooz Khan and Mukesh Nitinwriting of the manuscript; Pinki Raj Sahu and Sanyukta Kumar helped with proof reading of the manuscript; Scientist Mukesh Nitin helped with designing, guiding, manuscript and proof reading of the research article.
Acknowledgement: I would like to acknowledge the Head, Department of Tech. Biosciences, Digianalix for providing research facilities and guidance in this project work.
Conflict of interest: There is no conflict of interest in the presentation of this paper.
Funding source: There is no funding source.
