1Department of Chemistry, Illinois State University, Normal, IL 61790, USA.
2SGX Medical LLC, Bloomington, IL 61704, USA.
Marjorie A Jones
Tel: +1-309-438-2366;
Email: majone3@ilstu.edu
Received : Sep 04, 2023 Accepted : Oct 17, 2023 Published : Oct 24, 2023 Archived : www.meddiscoveries.org
Brain cancers, such as Glioblastoma Multiforme (GBM), exploit the body’s own immune system to support tumor growth within their microenvironment. GBM, an incurable cancer known for its highly invasive characteristics, utilizes various pro-inflammatory pathways to mediate the body’s response, one of them being the CCR5 signaling pathway. Through cell viability assays, fluorescence microscopy and spectroscopy, we discovered that the small molecule CCR5 antagonist Maraviroc (MVR), which has already been found to inhibit the invasion of primary GBM cells in vitro, has no significant effect on the cell viability or apparent cell structure of C6 glioma or PC-12 neuronal cells in vitro. However, PC-12 cells appear to exhibit a hormetic response in viability as a function of MVR concentration. In fluorescence experiments with a nitric oxide (NO) probe, we also found that MVR causes no changes in intracellular NO levels in C6 or PC-12 cells. Our research findings contribute to the ongoing study of the potential repositioning of MVR into therapeutics for brain cancers and other neurological conditions.
Keywords: CCR5; Maraviroc; Glioblastoma; C6 glioma cells; PC-12 neuronal cells; Cell viability; Nitric oxide; Fluorescence.
Glioblastoma Multiforme (GBM) is a highly aggressive type of brain cancer in adults that has a very poor prognosis. Despite advances in treatment, the average survival time for patients with GBM is still less than two years [1]. The ability of GBM to invade surrounding brain tissue is one reason why the disease remains incurable to this day [2]. Complete surgical removal is very difficult, and chemoradiotherapy is commonly used for treatment even after surgery [1].
The CCR5 pathway is a pro-inflammatory signaling pathway in the immune system. CCR5 (chemokine receptor type 5) is primarily expressed on the surface of many immune cells, and when it is activated by chemokines such as CCL3, CCL4 and CCL5, the receptor triggers a signaling cascade that leads to recruitment of immune cells to sites of infection or inflammation to help eliminate pathogens and promote tissue repair [3]. However, excessive activation of the CCR5 pathway can be detrimental, as it can lead to chronic inflammation, tissue damage, neuronal death, and cancer metastasis [4]. Inhibiting this pathway, by methods such as neuronal knockdown of CCR5, ligand knockout, and CCR5 antagonist treatment, has shown promising results in reducing inflammation and improving tissue repair and locomotor recovery in preclinical models of Traumatic Brain Injury (TBI) and Spinal Cord Injury (SCI) [5,6]. The antagonist used in these studies, CCR5 small molecule inhibitor maraviroc (MVR), originally used in HIV therapeutics due to CCR5’s role as an HIV coreceptor, is particularly of high interest as it is considered to be a new therapeutic strategy for various neuroinflammatory diseases [7]. In a recent study on MVR’s effects after TBI in mice, it was demonstrated that MVR decreases neuronal apoptosis by inhibiting neurotoxic reactive astrocyte activation while also reducing the neuroinflammatory response by decreasing the expression of inducible Nitric Oxide Synthase (iNOS) [8].
n the context of brain cancer, reactive astrocytes are known to contribute to the development and progression of GBM and other treatment-resistant glioma using ion channels and transporters [9]. Nitric oxide (NO) is a small and short-lived pleiotropic messenger molecule that plays a critical role in multiple cellular processes. It plays a complex role in the pathophysiology of GBM, as it maintains the balance between tumor progression and suppression [10]. It has been found, histochemically, that there is increased expression of nitric oxide synthase (NOS) in viable tumor cells [11]. NO is known to regulate vasodilation and vascular permeability, which may contribute to tumor cell survival [10]. Low levels of NO are associated with tumor cell growth, angiogenesis, and invasion, while high levels of NO can induce apoptosis and inhibit angiogenesis [12]. Since the delicate balance of NO levels is crucial in the human body, examining the intracellular levels of NO with and without MVR treatment in our in vitro cell models was considered of high interest for our research.
When under stress, both glial cells and neurons can contribute to increased NO production. Astrocytes generate NO when inducible Nitric Oxide Synthase (iNOS) is activated, but this is primarily during stress and inflammatory conditions because astrocytes have low levels of neuronal Nitric Oxide Synthase (nNOS), which is responsible for the general release of NO for physiological processes [13]. On the other hand, since neurons express more nNOS than astrocytes, they are reported to produce more NO in normal conditions [14]. With this current knowledge, it is assumed that neurons produce more total nitric oxide than astrocytes under stressful conditions due to the already existent intracellular NO produced by neurons. With axenic neuronal and glial cell model systems, we can compare NO levels in different conditions between the two cell types to evaluate this hypothesis.
Given the widespread presence of CCR5 in various tumor types, clinical trials are already underway to explore the potential therapeutic repositioning of MVR [4]. A study has found that MVR inhibits the invasion of primary GBM cells, demonstrating that it could be a potential therapeutic agent for slowing the progression of cancer in the brain [15].
To study GBM and develop new agents for treatment, primary human and animal cultures are important since they are directly derived from tumor tissue, but due to the possible interference of other cell types and microorganisms in these cultures, it is difficult to evaluate an agent’s direct effects on a cell-to-cell basis, thus axenic cell model systems also have importance. With axenic cultures, the mechanism of action by which MVR works can be further investigated. By examining MVR’s effects on neurons and glial cells individually, the specific responses and pathways triggered in those cells by MVR can be better understood.
Among the many cell lines used for glial cells, C6 glioma, a rat cancer cell line, has been the most common model for neurooncology studies [16]. With similar gene expression to human gliomas as well as high rates of tumor growth, invasion, migration, and angiogenesis, C6 glioma is considered to best simulate the behavior of human GBM [17]. Culturing of this popular glioma model allows for extensive research on the tumoricidal activity of drugs in relation to their activity against primary GBM cell lines.
In neurobiology research, the PC-12 cell line has been the preferred neuronal cell model due to its favorable neurogenic characteristics and morphology [18]. We report here our work with both cell types in vitro to explore other potential direct effects of MVR. By examining both cell model systems, we aim to contribute to a comprehensive understanding of the effects of drug treatment. Our investigation holds particular significance as it provides insight on how neural tissue may respond to these interventions. Furthermore, the utilization of a dual model system in our studies makes our findings more relevant and applicable. Therefore, our research helps advance our current understanding of drug response, serving as a foundation for further exploration in animal models and clinical studies.
From examining cell viability and intracellular NO levels in this study, we provide a model system that can be used to obtain valuable insights into the potential use of Maraviroc (MVR) as a therapeutic option for GBM treatment as well as other neurodegenerative conditions and neurological diseases
Cell cultures
From the American Type Culture Collection, Manassas, VA, USA (ATCC CCL-107), axenic Rattus norvegicus C6 glioma cells were purchased and cultured in sterile 6-well plates (GIBCO; Waltham, MA, USA) with low-glucose, no phenol Dulbecco’s Modified Eagle’s medium (DMEM; Sigma Life Sciences D6429; St. Louis, MO, USA) supplemented with 1.5% (v/v) heat-inactivated fetal bovine serum (GIBCO; Waltham, MA, USA), 10% (v/v) horse serum (ATCC; Manassas, VA, USA), and 4% L-glutamine following the method of Platt et al. (2022) [19]. This medium supplemented with sera was referred to as “complete medium”, while the medium without any supplementation was designated as “incomplete”.
Axenic PC-12 Pheochromocytoma cells (ATCC- CCL 1721.1), purchased from ATTC, were cultured in the same complete medium used for the C6 glioma cells. PC-12 cells were grown in separate sterile 6-well plates pre-coated with sterile Poly-Llysine (Sigma-Aldrich; St. Louis, MO, USA).
All cells were grown in an incubator set at 37OC with 5% CO2 and >95% humidity. To harvest, and transfer the cells, trypsin was used, following the instructions provided by the manufacturer. The trypsin was then promptly neutralized by adding complete medium, which contains α-1 antitrypsin. Resultant cell preparations were then centrifuged (Labnet Hermle Z 400K, Edison, NJ, USA) at 1000 rpm for 10 minutes at 7OC. The supernatant was removed before resuspending the cell pellets in complete medium and plated as needed.
For experimental procedures, cells were grown to confluency before being harvested as previously described, resuspended in 1 mL of complete medium, diluted ten-fold, and then counted with a Scepter 2.0 Handheld Automated Cell Counter (Millipore, Burlington, MA, USA) with a 60 µm sensor. Cells were subsequently seeded into either 96-well or 24-well sterile cell culture plates as appropriate for each experiment. Maraviroc (MVR) was introduced 24 hours after cell plating. Cell viability assays, microscopy, and fluorescence measurements for nitric oxide were performed when the cells reached confluency in the cell culture plates, on the second or third day of culture.
Cell viability assays
To determine the cytotoxicity and dose-dependent effect of maraviroc (Millipore PZ0002, USA) on our cell model systems, cell viability was determined using the 3-[4,5-dimethylthiazol2-yl]-2,5-diphenyltetrazolium bromide (MTT) assay following the procedure of Mosmann (1983) [20]. C6 glioma or PC-12 cell suspensions were seeded into each well of separate Poly-Llysine coated, sterile 96-well plates (6000 cells/well) and left to grow for 24 hours. Thereafter, MVR at different concentrations were added to the wells. Final MVR concentrations in the wells ranged from 10-80 µM. Then cell viability was evaluated.
MVR stock solutions were prepared in Dimethyl Sulfoxide (DMSO) before being diluted into complete medium with a final DMSO concentration of 1% (v/v). The experimental blank contained 1% DMSO, and complete medium without cells. Negative control wells contained 1% DMSO, and complete medium with cells. After introducing MVR or control, cells were incubated for 24 hours. After incubation, the cells were in their second day of culture and had reached confluency.
Experiments proceeded by replacing the complete medium containing DMSO/MVR with 100 µL of incomplete medium. Ten µL of MTT reagent (5 mg/mL water) were then added to each well. Cell culture plates were incubated for 60 minutes before 100 µL of solubilization solution (10% v/v Triton X-100, 1 mM HCl in isopropanol) was added to lyse the cells and dissolve the formed formazan crystals. Absorbance values were measured at a wavelength of 595 nm using a Bio-Rad® Microplate Reader Benchmark. All control and experimental groups consisted of n=5 replicates. Absorbance values from each group were corrected with the blank’s mean absorbance and converted to represent percent of the negative control. Results are reported as mean ± standard deviation. Differences in values between control and treatment groups were evaluated for statistical significance using one-way Analysis Of Variance (ANOVA) and the Tukey post hoc test with a significance level of α <0.05. Statistical significance is symbolized by an asterisk in the figures shown in the Results Section.
Cell culture and fluorescent probe preparation for analysis of nitric oxide levels
To examine the effects of MVR on intracellular NO levels, sterile 24-well cell culture plates were used. Cell suspensions were seeded into each well to a total volume of 270 µL (12000 cells/well). The plates were left in the incubator for 24 hours to allow the cells to grow.
The following day, 30 µL of MVR, already diluted into complete medium as described previously, were added to wells to reach a final MVR concentration of 80 µM. All wells were treated with the same concentration of MVR and had a final DMSO concentration of 1% (v/v). Control wells had only 1% DMSO. After compound addition, cells were incubated for another 24 hours.
Intracellular NO was detected using a fluorescent probe. 2’-7’-dichlorofluorescin diacetate (DAF-FM; excitation/emission: 495/515 nm) was purchased (EDM Millipore Corp., Burlington, MA, USA) and prepared in accordance with the manufacturer’s instructions. In the presence of intracellular NO, DAFFM fluoresces and can therefore be qualitatively and quantitatively analyzed using fluorescence microscopy and fluorometry, respectively. Before adding this NO probe, the medium was removed from all wells in the cell culture plates. DAF-FM was diluted with incomplete medium to a concentration of 10 µM. Then, 270 µL of this diluted DAF-FM solution were added to all wells then incubated for 60 minutes before microscopy and measurements were performed.
Fluorescence microscopy and fluorometry measurements
Following incubation, medium was discarded from all wells. Cells were washed with sterile 37OC Phosphate Buffered Saline (PBS) before a final 500 µL of warm PBS was added to all wells. Using an epifluorescence microscope (Keyence BZ-X810, Itasca, IL, USA) with a TRIT-C filter cube, bright field and fluorescence microscopy was performed at 20 magnification. By imaging control cell wells (1% DMSO) first, scope recording values, light exposure, and contrast settings were set to levels appropriate for optimal fluorescence acquisition. The imaging conditions used for the control were utilized for all the subsequent images. Bright field and TRIT-C (thus NO positive) fluorescent images were taken for all wells and then overlaid using Keyence analyzer software and cells with red color are positive for NO. Higher intensity of DAF-FM red fluorescence indicates higher levels of intracellular NO.
After capturing all images, fluorescence spectroscopy measurements were obtained. The PBS was removed and replaced with 200 µL of 1% (w/v) Sodium Dodecyl Sulfate (SDS) in water to solubilize the adherent cells. Samples from individual wells for each condition were pooled and then pipetted into a fluorescence cuvette to be measured by a fluorescence spectrometer (LS-55, Perkin-Elmer; Waltham, MA, USA). DAF-FM probe excitation and emission wavelengths were set, and fluorescence intensity was recorded every second for 60 seconds.
Maraviroc dose-dependent effect on C6 and PC-12 Cell viability
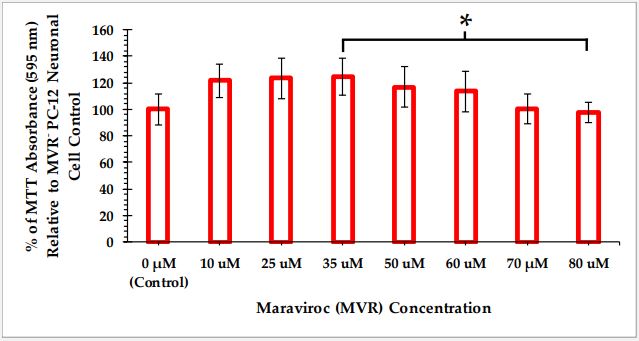
The average absorbance at 595 nm of five replicate wells for each concentration of MVR is reported as percent relative to the cell only negative control’s average absorbance. MTT cell viability data of C6 glioma cells and PC-12 neuronal cells as a function of MVR at different concentrations are reported in Figures 1 and 2, respectively.
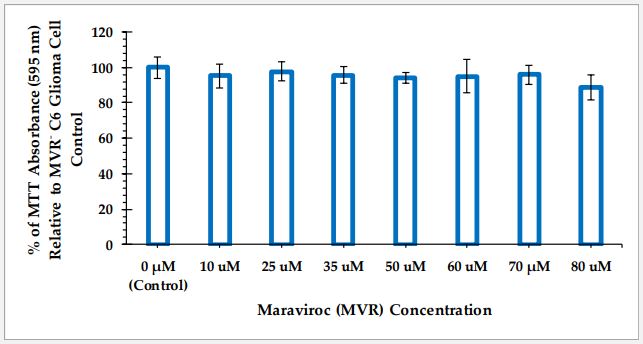
To compare the effect of the different MVR concentrations on the cell viability of C6 glioma cells, a one-way ANOVA was performed on the data presented in Figure 1. The omnibus test results show that there are no statistically significant differences in the cell viability between any of the MVR treatment groups (F (7,32) = [1.347], p=0.26).
One-way ANOVA performed on the PC-12 cell viability data displayed in Figure 2 reveals that there is a statistically significant difference in the cell viability between at least two of the MVR treatment groups (F(7,32) = [3.736], p=0.005). Tukey’s HSD test for multiple comparisons found that the cell viability is significantly different between PC-12 cells treated with 35 µM MVR and those treated with 80 µM MVR (p=0.049, 95% C.I. = [0.107, 53.693]). These data suggest a hormesis effect of MVR on this cell type. Cell viability after all MVR treatments, however, are not significantly different than that of the control.
Maraviroc effect on C6 and PC-12 morphology and intracellular nitric oxide levels
Brightfield and fluorescent images were obtained for all conditions and are displayed in Figure 3 for C6 cells and Figure 4 for PC-12 cells. Treatment of 80 µM MVR caused no change in the morphology of both cell types. Compared to their respective controls, C6 and PC-12 cells treated with MVR remained adherent, membranes stayed intact, and the presence of cell processes was unaffected (Figure 3, A & B; Figure 4, A & B).
Regarding Nitric Oxide (NO) fluorescence levels, fluorescent images C & D in Figures 3 and 4 demonstrate that few, if any, of either cell type did not exhibit the characteristic NO fluorescence. Also, there are no clear observable differences in intracellular NO levels between control cells and those treated with MVR for both cell types. Therefore, fluorometry was necessary to quantitatively ensure that there are no differences in fluorescence intensities.
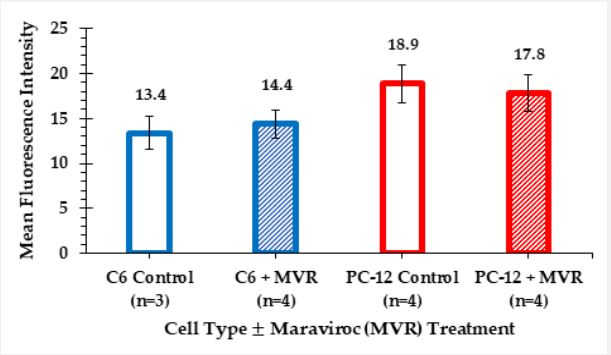
Our fluorescence spectroscopy measurements further suggest that MVR has no effect on intracellular NO levels of the pooled samples for either cell type (Figure 5). There is no clear difference between the fluorescence intensities of the control group and the MVR treated group within each cell type. However, PC-12 neuronal cells do have modestly higher NO signal, with and without MVR treatment, compared to C6 glioma cells.


From our MTT cell viability experiments, it is evident that maraviroc at concentrations up to 80 µM does not affect the cell viability of either C6 glioma cells or PC-12 neuronal cells. However, PC-12 cells seem to be more sensitive to maraviroc compared to C6 cells. This apparent hormesis effect is of interest and may have important implications for MVR therapeutic use. The absence of any impact on C6 cell viability by MVR aligns with the findings reported by Novak, et al. [15], who demonstrated that MVR did not affect the viability of primary glioblastoma cells.
Interestingly, when comparing the fluorescence spectroscopy data between our cell model systems, a modest (20-30%) difference was found in intracellular NO levels between C6 and PC12 cells while the fluorescence microscopy data do not reveal these modest differences. Our results support other reports that neurons produce more NO than astrocytes (a type of glial cell) in all conditions [14], stressful or not. However, additional studies are needed to investigate this difference.
Our brightfield microscopy images show that MVR treatment causes no observable changes in the morphology of either C6 or PC-12 cells. When MVR was introduced to cells incubated with a NO probe, fluorescence levels did not change from that of the no MVR controls, as shown both visually in our microscopic image overlays and fluorescence spectroscopy measurements. Intracellular NO levels did not change after MVR addition. These results extend the work of Liu, et al. [8] who reported that addition of maraviroc in a mouse model of brain injury decreased the expression level of inducible nitric oxide synthase and promoted animal recovery. Our results imply that other isozymes of nitric oxide synthase still present in cells remain functional. Future work should evaluate if MVR has any direct effect on nitric oxide synthase isozymes.
Strengths of our study include the use of axenic cell cultures, as model systems, as these models allowed us to evaluate MVR’s individual effects on neuronal cells versus glioma cells. However, we are limited in our ability to assess the roles of each of these cell types in the pathology of GBM and other human gliomas. Since we have used only axenic cell cultures, our experiments lack the complexity of the in vivo environment. This limits our findings from being able to thoroughly demonstrate the response of whole organisms. With our focus specifically on MTT cell viability and intracellular NO levels, our study is not able to demonstrate all interactions and signaling pathways that contribute to responses to maraviroc. Therefore, additional studies with other cell types or co-cultures can help better represent the diversity of cell populations in the brain or the tumor microenvironment. Future work evaluating the effects of MVR on multiple GBM cell lines would be useful and confirm our findings. Coupling MVR with other forms of treatment is also of interest as a potential continuation of this study. Research on the extracellular NO levels following MVR treatment would further add to a comprehensive understanding of NO signaling changes caused by MVR. Our current results contribute to an improved understanding of MVR as a new therapeutic option for treating GBM and other neurological conditions.
Our results suggest that maraviroc is not toxic in vitro to both cell models which supports the results by others [5,6,8] who have used CCR5 inhibition by MVR and other methods, such as CCL3 ligand knockout, in preclinical models of traumatic brain injury and spinal cord injury. Our findings suggest that MVR may not only be used as a treatment after traumatic injuries but also for brain cancers like GBM. Thus, MVR may be utilized safely in animal models and clinical studies, but more testing with other conditions is necessary to evaluate the safety of repositioning this drug for potential therapeutics for brain cancer, neurotrauma, and other neurological diseases.
Author contributions: “Conceptualization, S.M.Z, D.C.P, and M.A.J.; methodology, D.C.P, and M.A.J.; formal analysis, S.M.Z, D.C.P, and M.A.J; investigation, S.M.Z, D.C.P, and M.A.J.; resources, M.A.J; writing—review and editing, all authors; All authors have read and agreed to the published version of the manuscript.
Funding: This research received no external funding.
Institutional review board statement: Not applicable.
Informed consent statement: Not applicable.
Data availability statement: Data used are all presented in the manuscript.
Acknowledgments: S.M.Z was supported by an Undergraduate Antonini Fellowship, ISU.
Conflicts of interest: The authors declare no conflict of interest.
