1Scientific Director, Centre for Human Reproduction, Scientific Director cum Owner Dr Kulvinder Kaur Centre for Human Reproduction, India.
2Scientific Director, Ex-Rotunda-A Centre for Human Reproduction, India.
Kulvinder Kochar Kaur
Tel: 91-9501358180, 91-181-4613422, Fax: 91-181-4613422;
Email: kulvinder.dr@gmail.com
Received : May 30, 2025 Accepted : Jun 17, 2025 Published : Jun 24, 2025 Archived : www.meddiscoveries.org
Diabetic Kidney Disease (DKD), represents the maximum frequent microvascular complications of Type 2 Diabetes Mellitus (T2DM), that is the greatest etiological factor of End Stage Renal Disease (ESRD). Despite precise mechanistic modes of even now DKD continues to be uncharted, programmed cell death has been taking in the phenomenon along with generation of diabetic kidney damage, inclusive of ferroptosis. Ferroptosis portrays a kind of cell demise, where Reactive Oxygen Species (ROS) along with lipid peroxides get generated by iron metabolism as well as their accrual for generating fatal toxicity in view of cells are not capable of metabolizing them in a smooth manner. It has been isolated to possess a crucial part in the generation of therapeutic reactions to plethora of kidney diseases for instance Acute Kidney Injury (AKI), renal cell carcinoma, polycystic kidneys in addition to DKD. In the last 4-5 years substantial evaluation of ferroptosis has been performed in diabetic subjects along with in animal models, nevertheless, precise mechanistic modes have not been displayed. Here we have provided a comprehensive narrative review regarding the controlling of mechanistic modes of ferroptosis. Additionally, we have detailed the observations that corroborate the association of DKD with ferroptosis as well as described how ferroptosis might work in the form of a plausible target for generating innovative therapeutic strategies for DKD therapy for furthering scientific researchers to work for unraveling the pathogenesis of DKD for improvement of clinical outcomes of DKD patients.
Keywords: Diabetic kidney disease (DKD); Ferroptosis; Iron metabolism; Lipid peroxides accrual; Reactive oxygen species (ROS); Cell death.
In the last 2 decades the incidence of Diabetic Kidney Disease (DKD) has escalated with the escalating prevalence of Type 2 Diabetes Mellitus (T2DM) [1]. In the form of the maximum frequent microvascular complications of DM, DKD possesses bad prognosis in addition to restricted treatment methodologies. As per the epidemiological statistics, DKD represents the basic exposition for world over End Stage Renal Disease (ESRD) [2]. Currently, the availability of clinical treatment modalities is inclusive of Angiotensin Converting Enzyme (ACEIs), A angiotensin II Receptor Antagonists (ARBs), Sodium–G Lucose Cotransporter 2 (SGLT2)- hampering agents [3,4]. Despite, such treatment modalities possess the capacity of diminishing the propagation of DKD to certain magnitude, the incidence of DKD has continued to be substantially greater in recent decades with robust sequelae along with adding cost prohibitive load. Variable studies have corroborated that plethora of pathophysiological factor abnormalities aid in the initiation as well as generation of DKD, for instance i) Haemodynamic aberrations, ii) Oxidative stress (OS) in addition to iii) Cell demise, iv) Genetic along with epigenetic controlling [4-6]. Dependent on the complex pathogenetic mechanistic modes underlying as well as no ideal treatment results of DKD, estimating the precise pathogenesis in addition to generating correct treatment modalities is imperative. Nevertheless, escalating corroboration has taken into account that the combination of plethora of factors along with cell demise evokes considerable interest as well as is believed to be a direct factor influencing DKD [7]. Recently ferroptosis has been illustrated to possess a critical part in the initiation as well as propagation of DKD [7].
Dixon et al. [8], in their pioneer work initial posited in addition to validated its presence in cancer stimulated by erastin [8]. Apart from other kinds of non-apoptotic cell demise, ferroptosis based on the iron overload, have the properties of accumulation of lipid peroxides [8,9]. Subsequent to the development of ferroptosis, unique alterations are witnessed in the mitochondrial morphology, inclusive of diminished/even elimination of cristae, rupturing of outer membrane, an escalated quantity of intracellular ROS, which get followed by rupturing of cell membrane resulting in cell demise. As per the biochemical changes intracellular accrual of iron, enrichment of Reactive Oxygen Species (ROS) in addition to accumulation of lipid peroxides presentation takes place at the time of circumstances of ferroptosis [10]. Once further advancements of scientific research occurred invention of ferroptosis proved to be crucial in plethora of variety of diseases inclusive of cancer, neurological diseases, Cardiovascular Disease (CVD) along with kidney diseases [11]. In reference to kidney diseases maximum researchers have basically concentrated on the association of ferroptosis as well as Acute Kidney Injury (AKI), renal cell carcinoma, in addition to polycystic kidneys [11], along with germanely minute validation to embrace the degree by which ferroptosis aid in generation of DKD. Previously we reviewed articles on pathophysiology, therapy of DKD inclusive of role of epigenetics in DKD, ferroptosis in Breast Cancer (BC), in NAFLD as well as NAC in Polycystic Ovary Syndrome (PCOS) & AKI [12-21]. The objective of this narrative review was to investigate role of ferroptosis in DKD which aid in generating targeted therapies against ferroptosis for the treatment of DKD in future.
Here we conducted a narrative review utilizing search engine pubmed, google scholar; web of science; embase; Cochrane review library utilizing the MeSH terms like DKD; Epigenetics; ferroptosis; Glutathione Peroxidase 4 (GPX4); lipid peroxidation; Divalent Metal Transporter (DMT); Ferritin; Oxidative Stress (OS); Ferritinophagy; AMPK; Nuclear factor erythroid-2-Related Factor-2((Nrf2)/ Kelch-like-Epichlorohydrin (ECH)-associated protein 1 (KEAP1); HMGB1; ZIP14; NAC; Herbal products; curcumin analogs; Apabetalone sodium butyrate from 1998 to 2025 till date.
We found a total of 900 articles out of which we selected 158 articles for this review. No meta-analysis was done.
Crucial mechanistic modes of ferroptosis
In the last 20 yrs the insight responsible for the mechanistic modes of ferroptosis signaling has undergone remarkable propagation. To start with invention of role of cysteine - Glutathione (GSH), in addition to Glutathione Peroxidase 4 (GPX4), in hampering of ferroptosis in the implementation of lipid peroxidation has been corroborated. Additionally, GPX4 autonomous ferroptosis surveillance mechanistic modes have been displayed. Herein we detail the main mechanistic modes of ferroptosis (Figure 1) [22].
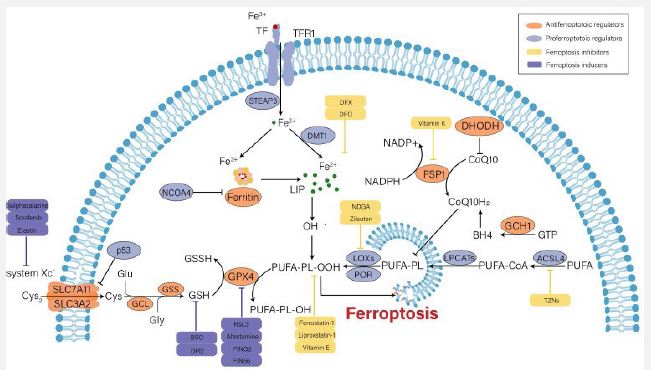
Courtesy reference no-22- Iron metabolism disorder, lipid peroxidation and decreased antioxidant capacity are involved in the occurrence of ferroptosis, and several key regulators (e.g., system Xc-, GPX4, p53 and ACSL4) play an important role in monitoring ferroptosis. Some recently developed compounds can induce or inhibit ferroptosis by targeting these key regulators.
Iron metabolism aberration
In case of normal physiological situations, binding of circulating ferric iron (Fe3+) takes place to Transferrin (TF), whose transportation takes place into cells by the membrane protein Transferrin Receptor 1 (TfR1). Following that reduction of Fe3+ takes place to Fe2+, by Six Transmembrane Epithelial Antigen of Prostate 3 (STEAP3). Divalent Metal Transporter (DMT) controls the liberation of Fe2+, into the Labile Iron Pool (LIP) for more use, along with the storage of the remaining Fe2+, in the form of ferritin. gets constituted by Ferritin Light Polypeptide (FTL1) as well as Ferritin Heavy polypeptide (FTH1), in addition to possesses the capability of catalyzing the transformation of Fe2+, into Fe3+ for the sustenance of iron homeostasis in the cytoplasm [23]. On the one side escalating free iron had the capacity of generating free radicals for instance hydroxyl radical (OH)-* that cross talks with Hydrogenperoxide (H2O2). Oxidative Stress (OS) results from the dysequilibrium of rates of generation of free radicals along with their detoxification, eventually stimulating ferroptosis through lipid peroxidation [24]. Conversely, extra Fe2+, stimulate the activation of ferric enzymes for instance Lipooxygenases (LOX) as well as potentiates lipid peroxidation in the cytomembrane, which ultimately results in cell demise [25]. Thereby, escalated iron uptake in addition to diminished iron extrusion would escalate the susceptibility of the cells to oxidative injury along with ferroptosis. Ferritinophagy by definition portrays the selective autophagic breakdown of ferritin resulting in accumulation of cytosolic Fe2+. Nuclear receptor Coactivator 4 (NCOA4) represent the critical controller of ferritinophagy [26]. In case of Renal Tubular Epithelial Cells (RTEC) Knock Out (KO) of NCOA4 possesses the capability of attenuating ferroptosis which takes place subsequent to autophagy [27].
Accrual of lipid peroxides
Lipid peroxidation is believed to be a start signal in reference to ferroptosis [28]. In contrast to unsaturated fatty acids along with monounsaturated fatty acids, Polyunsaturated Fatty Acids (PUFA), possess greater susceptibility to lipid peroxidation, in view of greater instability of carbon- carbon double bonds, that aid in ferroptosis [29]. Esterification of the free PUFAs takes place by activation of Acyl-CoA Synthetase Long-chain family member 4 (ACSL4) as well as transported to membrane Phospholipids (PL’s) by the Lysophosphatidylcholine Acyltransferase 3 (LPCAT3).
Subsequenly oxidation takes place to toxic lipid peroxides by LOXs. Out of the PUFAs associated PL’s, Phosphatidylethanolamine (PE) of Arachidonic Acid (AA) as well as Adrenic Acid (ADA) constitute the basic substrates at the time of events of lipid peroxidation [24]. The robustness of ferroptosisis based on the magnitude of the accumulation of PUFAs. Furthermore, the actions of ACSL4 in addition to LOXs are completely requisite for the accomplishment of ferroptosis. Doll et al. [30], posited that ACSL4 driven ferroptotic cell demise takes place by remodeling of intracellular lipid constitution [30]. Hampering agents of LOXs for instance liproxstatin -1 along with Vitamin E prove to be efficacious in the avoidance of ferroptosis [31]. As per the invention of recent studies cytochrome P450 Oxidoreductase (POR) was responsible in guiding lipid peroxidation [32,33]. Broader expression of POR has been observed in Endoplasmic Reticulum (ER) in lipid peroxidation, that passes electron from Nicotinamide Adenine Nucleotide Phosphate (NADPH) to Cytochrome P450s ((CYPs) as well as other proteins [34]. Lipid peroxidation might be started by POR by promoting Fenton reactions in the heme constituent of CYPs [32]. Nonetheless, one extra study illustrated that the crosstalk amongst POR in addition to its downstream electron receptors were not needed for the ferroptosis being stimulated, along with POR by itself was capable of autonomously resulting in dysfunctional PUFA possessing phospholipids membrane [33]. Hence, the part of the POR in ferroptosis continue to be uncharted, and greater evaluation is required.
Dysequilibrium of antioxidant capability
In case of normal physiological situations, intracellular Glutamate (Glu) gets exchanged with the extracellular Cysteine (Cys2) in a 1:1 ratio via the cysteine- glutamate transporter receptor – (System Xc). System Xc represents a disulfide heterodimer, that comprises of a light chain subunit -solute carrier family generation 7 members 11 (SLC7A11), along with a heavy chain subunit (SLC3A2) chain that has substantial placement on the cell membrane [10]. Subsequent, to entry in the cell reduction of Cys2 to cysteine (Cys) takes place for the GSH generation, that gets catalyzed by the Glutamate -Cysteine Ligase (GCL) as well as Glutathione Synthetase (GSS) [9]. GSH portrays an intracellular antioxidant which takes part in enzymatic in addition to nonenzymatic reactions for the sustenance of normal quantities of H2O2 in the cells. GPX4 possesses the capacity of reducing lipid hydrogen peroxide (L-OOH), into Lipid Hydroxyl derivative (L-OH), that is not toxic in GSH based way, for stopping ferroptotic cell demise by inimical lipidperoxides [35]. Thereby, intracellular GSH elimination/ inactivation of GPX4 stimulated by variable factors would diminish the clearance rate of toxic lipid peroxides along with lead to ferroptosis. For instance, erastin possesses the capability of hampering cysteine uptake as well as GSH formation by blockade of System Xc, therefore promoting ferroptosis [10]. RSL3, stimulates ferroptosis by inactivating GPX4actions [36]. Additionally, it got corroborated that inducible KO of GPX4 was capable of stimulating acute kidney failure via the ferroptotic pathway [37].
Undeniably GPX4 represents a crucial controller of ferroptosis. Nevertheless, three pathways sovereign of GPX4 have been isolated in the last decade inclusive of i) Ferroptosis Suppressor Protein 1 (FSP1)/ ii) Ubiquinone (alias coenzyme Q10 (CoQ10)), iii) GTPcyclohydrolase1(GCH1) along with, iv) Dihydroorotate Dehydrogenase (DHODH). FSP1 possesses the capability of regenerating the reduced form of CoQ10 alias Ubiquinol (CoQ10H2) with the use of NADPH. Ultimately, CoQ10 depletes lipidperoxides via an oxidation reaction [38]. GCH1 represses ferroptosis basically by forming lipophilic antioxidant tetrahydrobiopterin (BH4) as well as remodeling of lipid membrane milieu for escalating CoQ10H2 in addition to eliminating PUFAs [39]. Furthermore, DHODH has been observed to hamper ferroptosis by reducing CoQ10 [40].
Other controllers
Apart from canonical controlling pathways manipulation of ferroptosis takes place in a plethora of manners for instance i)p53 ii)Nuclear factor erythroid-2-Related Factor-2((Nrf2) along with iii)AMP-Activated Protein Kinase (AMPK) [41,42], of which maximum attractive has been p53 in view of its debatable part in ferroptosis.
i) p53 possesses double part in ferroptosis [41]. It got displayed that p53 was capable of facilitating ferroptosis by repressing the expression of SLC7A11 in transcriptional as well as non-transcriptional ways, that resulted in diminished uptake of Cys2 in addition to led to dysfunctional GSH biogeneration [44]. Furthermore, p53 possesses the capability of transactivation, spermidine/spermine acetyltransferase (SAT1), that portrays a rate restricting enzyme in case of polyamine metabolism. SAT1 was illustrated to stimulate ferroptosis by increasing the expression of arachidonate 15 lipooxygenase (ALOX15) [44]. Chu etal. [45], found that ALOX12 manipulated lipid peroxidation was involved in p53 based ferroptotic reactions [43]. Additionally, Glutaminase (GSP2), Cytochrome-c Oxidase (sCO2) along with variable noncoding RNAs got validated in the form of target molecules of p53 manipulated ferroptosis [46]. Nonetheless, p53 apparently hampered ferroptosis in some cellular metabolic milieu. Xie et al. [47], in a study corroborated that p53 was capable of binding to Dipeptidyl Peptidase -4 (DPP-4) for diminishing the DPP-4based oxidation of PUFAs for the hampering of ferroptosis in case of colorectal cancer [47]. Cyclin –based Kinase inhibitors1 (CDKN1A)/p21 upregulation by p53 aided in diminished sensitivity to ferroptosis in cancer cells. The mechanistic modes behind this are believed to be p21 stimulated escalated generation of GSH [48].
Advancements in pharmacology
Recognized that plethora of controlling factors are implicated in the generation of ferroptosis, targeting some pathways possesses the capability of stimulating or hampering events. Therapeutic applications of stimulating ferroptosis might be done for depleting cancer cells as well as repression of ferroptosis might work in the form of robust treatment of Cardiovascular Disease (CVD), neurodegenerative diseases in addition to DKD [10]. In the last decade small molecules have been corroborated to be of use in controlling ferroptosis (see Figure 2 for the evolution of ferroptosis hampering.
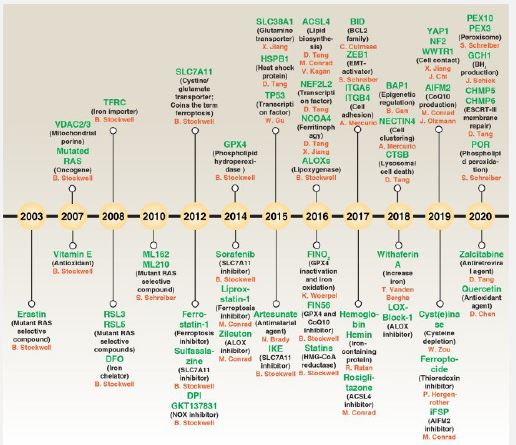
Ferroptosis controlling takes place in a myriad of manners- i) the cysteine import system System Xc- upstream of GPX4 gets hampered along with System Xc gets controlled by hampering agents for instance Imidazole Ketone Erastin (IKE), Sulfasalanazine as well as Sorafenib [49]. Additionally, ferroptosis induction is further feasible by direct controlling of the reduction of GSH, along with GPX4. GPX4 controlling might be done by hampering agents for instance, RSL3, ML162, ML-210, as well as FIN56, whereas GSH generation gets hampered by Butylthionine Sulphoamine (BSO) [50]. The crucial factor of ferroptosis, Fe (II)- might be hampered in a plethora of manners for instance by different hampering agents-i) Desferoxamine (DFO), Desferoxamine Mesylate (DFOM), 2, 2’Bipyridine (BP) in addition to Ciclopirox (CPX) [51].
Scientific research propagation of ferroptosis in DKD
Earlier models had observed direct correlation of iron overload along with proteinuria as well as tubular injury in patients with DKD in addition to Streptozocin (STZ) induced rat amongst model of type 1 diabetes mellitus [52]. Escalated iron quantities observed in the lysosomes of proximal tubular epithelial cells\had the capacity of stimulating renal impairment in DKD patients along with mice [53]. Feeding diabetic rats with diets containing lesser iron quantities / with ones possessing iron chelator possessed the capability of postponing the generation of DKD, pointing that iron overload was capable of accelerating kidney damage in DKD [54]. Zhao et al. [55], validated that the transferrin quantities as well as iron quantities in RTECs of DKD patients are greater, in contrast to healthy persons [55]. Collectively, these observations displayed that the formation of DKD is incapable of being separated from iron overload.
Currently a myriad of studies has corroborated that ferroptosis takes place in RTECs in case of diabetic animal models as well as stimulated inimicality of kidney working. Wang et al. [56], corroborated that ferroptosis was implicated in kidney damage in STZ stimulated type 1 diabetic mice in addition to db/db mice [56]. Subsequently, Kim et al. [57], observed that the quantities of SLC7A11, along with GPX4 displayed significant downregulation in renal biopsy samples, obtained from diabetic patients. In the meantime, their work further pointed that expression quantities of ferroptosis correlated biomarkers for instance iron quantities as well as lipid peroxides with toxicity were escalated in Transforming Growth Factor Beta1 (TGF-β1) stimulated NRK-52E cells in addition to type 1 diabetic mice [57]. Apart from RTECs, ferroptosis were capable of getting observed practically in all three kinds of renal cells in case of DKD. Zhang et al. [58], posited that amelioration of ferroptosis possesses the capability of avoidance of glucose stimulated podocytes damage [58]. Escalated fructose possesses the capability of stimulating podocytes ferroptosis along with eventually resulting in glomerular dysfunction [59]. In case of mesangial cells, molecules associated with ferroptosis get changed in the case of escalated glucose situations, that might get antagonized by DFO, a ferroptosis hampering agent [60]. Additionally, ferroptosis is the executioner in the event of endothelial cells impairment that results from hyperglycemia [61]. Therefore, escalating validation is emerging that ferroptosis actually stimulates the generation of DKD as well as hampering ferroptosis works in the form of therapeutic target in reference to DKD therapy.
The interactions of ferroptosis along with other cellular events in DKD
Escalating work in the field of ferroptosis in the context of variable diseases, invention of escalated mechanistic modes has been obtained [11,62]. Of greater significance, greater canonical controllers of DKD have been isolated for manipulating ferroptosis d efficaciously [57,63]. The aim here is to detail crosstalk amongst ferroptosis as well as the canonical pathogenesis of DKD (Figure 3).
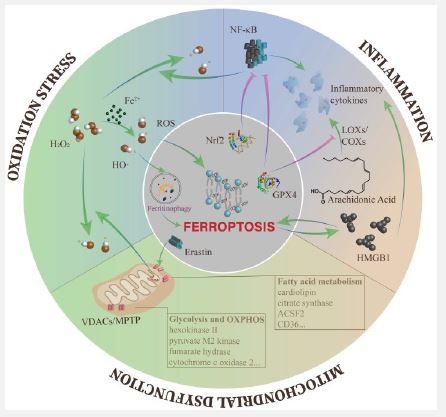
Courtesy reference no-22- The overproduction of ROS leads to different kinds of downstream impairments, including ferroptosis. Meanwhile, the Fenton reaction containing excess free iron is one of the main pathways to produce ROS. Ferroptosis and inflammation can affect each other and tend to form an auto amplification loop. In mitochondria, the opening of VDACs plays a key role in mitochondrial dysfunction during ferroptosis, and multiple metabolic enzymes are influenced by ferroptosis.
Ferroptosis along with oxidative stress: Pathogenesis of DKD is practically incapable of being disjoined from the overactivation of ROS, that portrays the communication area amongst metabolic aberrations in addition to haemodynamic alteration [64]. ROS accrual works out to disrupt each kind of kidney cells, at the level of gene, transcription along with protein, ultimately leading to inflammation, fibrosis as well as endothelial cells impairment [65].
In addition to the reduction of antioxidant capability, there is existence of a plethora of manner in the generation of ROS in DKD. Taken into account the surfeit iron in DKD, the Fenton reaction is an imperative part & parcel of generation of ROS [64]. With regards to ferroptosis the maximum direct pathogenic mechanistic modes resulting from ROS in lipid peroxidation eventually stimulates ferroptotic cell demise [66]. Additionally, ROS directly possess the capability of getting implicated in erastin stimulated ferroptosis in ferritin autophagy for the upregulation of accessibility of intracellular iron quantities [67]. Wang et al. [68], observed an intricate association amongst GPX4 in addition to DKD inclusive of urinary proteins, Scr, Estimated Glomerular Filtration Rate (eGFR) along with proportion of glomerulosclerosis observed in kidney specimens. Wang et al. [68], further posited that the GPX4 quantities in the tubulointerstitium worked in the form of an autonomous anticipator of results [68].
Collectively, ROS works in the form of intersection in reference to the effect of DKD stimulated ferroptosis. In view of that quercetin as well as Vitamin E are believed to be clinical treatments in reference to DKD patients, however such trials continue to be just in the animal model trials [69]. Recognized the actions of ROS in ferroptosis, possibility of generating efficacious anti DKD therapies for manipulating ferroptosis in DKD exists.
Ferroptosis along with inflammation: Accumulating corroboration has pointed significance of inflammation in the propagation of DKD [70]. Hyperglycemia stimulates the overexpression of chemokines as well as cytokines in the injured glomeruli in addition to renal tubules in case of DKD that accelerate renal damage via a myriad of mechanistic modes for instance i)Leukocyte infiltration ii)Degrading renal filtration barrier along with iii)Mesangial proliferation [70]. Such proinflammatory molecule further stimulate aforementioned damages via NFκB nuclear factor κB(NFκB), Janus Kinase / Signal Transcription and Transducer (JAK/STATs), TGF-β1 as well as SMA as well as Mad -related protein (SMAD) signaling pathways [71]. Negative controllers of ferroptosis are capable of hampering inflammation. AA portrays a precursor of bioactive proinflammatory manipulators. At the time of catalytic events, the cellular quantities of lipid peroxides directly influence the actions of enzymes Cyclooxygenases (COXs)-2 along with LOXs. Existence of possibility of blockade of LOXs activation is there in the cell once cell is in redox status for instance GPX4 activation [72]. Activation of GPX4 further possesses the capacity of diminishing ROS quantities by NFκB, therefore efficaciously diminishing cell injury stimulated by inflammation [73]. Additionally, Nuclear factor erythroid-2-Related Factor-2 ((Nrf2) possess the capability of hampering the transcription of NFκB [74], as well as proinflammatory genes getting encoded inclusive of Interleukin (IL-6), in addition to IL-1β [75]. ln case of an oxalate stimulated AKI mouse model, ferrostatin-1 attenuated neutrophils infiltration along with proinflammatory cytokines inclusive of C-X-C motif Chemokine Ligand (CXCL2) as well as IL-6 [76]. Actually, ferroptosis has the capacity of stimulating inflammation by liberation of IL-33 in addition to other uncharted pathways [9]. Ferroptotic cells are capable of liberating High Mobility Group Box1 (HMGB1) in an autophagy-based way. In the form of a Damage–Associated Molecular Patterns (DAMP), HMGB1 results in activation of immune cells for the liberation of plethora of chemokines along with cytokines [77]. On the other hand, myriad of proinflammatory cytokines Prostaglandin E2 (PGE2), IL-6), in addition to IL-1β have been corroborated to directly impact the ferroptosis initiation [62,78,]. HMGB1 further is capable of controlling ferroptosis in reaction to escalated glucose in case of DKD [60]. LOXs further have the capacity of activation of immune cells via LOXs proinflammatory metabolites inclusive of leukotriene B4 (LTB4), LTC4, LTD4, LTE4 as well as hydroxy eicosatetraenoic acid (18HETE) [79], that indirectly results in ferroptosis.
Accumulating corroboration pointed that ferroptosis is invariably associated with the presence of inflammation. Further interestingly, interactions amongst ferroptosis in addition to inflammation gets mutually stimulated, along with guides a local self-amplifying loop [80]. The internal association amongst ferroptosis along with aseptic inflammation is a point used by scientific researchers for unveiling association amongst ferroptosis as well as DKD.
Ferroptosis along with mitochondrial impairment: Mitochondria represent the crucial organelles involved in cellular energy metabolism [81]. Hyperglycemia causes metabolic substrate getting stored resulting in accrual of glycolytic intermediates as well as generation of mitochondrial ROS (mtROS), from the Electron Transport Chain (ETC), eventually stimulating mitochondrial impairment in addition to dysequilibrium of energy metabolism [82].
Mitochondrial homeostasis is critical for the sustenance of normal kidney working as well as the canonical morphological characteristics of ferroptosis is mitochondrial injury. Nevertheless, what determines the irreversible nature of mitochondrial injury at the time of ferroptosis? As per studies erastin along with its analogues have the capability of antagonizing the working of free tubulin present on Voltage Dependent Anion Channel 2/3(VDAC) in addition to VDACs opening. VDACs aid in controlling the passive diffusion of anionic hydrophilic mitochondrial metabolites [83]. Following that with the escalation of mitochondrial membrane potential, mitochondrial depolarization takes place with generation of extensive quantities of mtROS [84,85]. Additionally, glutamate possess the capability of escalating generation of mtROS via opening of Mitochondrial Permeability Transition Pore (MPTP) along with stimulating mitochondrial transmembrane potential draining as well as elimination of ATP [86]. VDACs opening in addition to generation of ROS result in mitochondrial Ca2+ overload, eventually stimulating mitochondrial impairment in addition to elimination of mitochondrial membrane cohesion [87].
Diminished glycolytic propagation was estimated in erastin stimulated ferroptosis as a result of escalated generation of ATP along with Oxidative Ohosphorylation (OXPHOS) [8,84]. Certain critical glycolytic enzymes were observed to be diminished in case of cancer cells that got RSL3treatment inclusive of Hexokinase II (HKII) as well as pyruvate M2 kinase [88]. Apart from variable enzymes implicated in mitochondrial respiration for instance sCO2(a protein essential for the assembly and function of cytochrome C Oxidase (COX), a crucial complex in the electron transport chain within the mitochondria) as well as Fumarate Hydrase (FH) [85]. Nonetheless, greater evaluating is the need for the aforementioned enzymes with regards to DKD models.
Furthermore, Fatty Acids (FA) metabolism in case of mitochondria is intricately correlated with lipid peroxidation in ferroptosis. Citrate synthetase as well as Acyl-CoA Synthetase Long-chain family member 2 (ACSL2) that control FA activation in addition to generation respectively, were isolated to be associated with erastin stimulated ferroptosis. Additionally, delivery of extensive quantities of mitochondria particular phospholipid Cardiolipin (CL) were done by oxidative GPX4-/- kidneys [30]. FA existence in the Tumor Microenvironment (TME) recently has been illustrated to stimulate CD8+T cells ferroptosis in a FA transporter (CD36) based way [89]. CD36 portrays the major pathway of FA uptake in addition to has considerable expression in full kidney cells [90]. RTEC cultured insubstantially greater glucose quantities in medium, observed escalated CD36 expression along with substantially greater FA uptake, which is CD36 modulated [91]. Therefore, there might be utility in exploring if CD36 is correlated with ferroptosis in DKD cases.
Ferroptosis associated controllers implicated in DKD
In case of DKD crucial controllers of ferroptosisis, implicated in canonical metabolic pathways displayed earlier have been evaluated widely. Therefore, here, we detail the mechanistic modes controlling ferroptosis in DKD is done.
Nrf2: Nrf2 represents a transcription factor of the basic- region leucine-zipper family [92]. Earlier studies have illustrated that Nrf2 possesses a critical part in reacting to antioxidative stress along with sustenance of redox homeostasis. In case of mechanistic modes of DKD, apart from Kelch-like-epichlorohydrin (ECH)-associated protein 1 (KEAP1)/ Nrf2)/ Antioxidant Response Element (ARE), Nrf2 is capable of hampering inflammation, for relief from kidney damage by repression of NFκB, Sirtuins (SIRT1), nucleotide-binding domain, leucine-rich-repeat containing family, pyrin domain-containing (NLRP3) inflammasome [74,93]. In a debatable fashion, Nrf2 apparently possesses double part in DKD. Zhao et al. [94], illustrated that Nrf2 insufficiency ameliorated kidney damage by upregulation of intrarenal ACE2 as well as, angiotensin 1-7 receptor in DKD [94]. Therefore, greater detailing Nrf2 part in DKD is done.
These days greater scientific research has been concentrated on the association amongst Nrf2 as well as ferroptosis. Nrf2 activating was initially observed to hamper ferroptosis by controlling the expression of NAD(P)H Quinone Oxidoreductase (NQO1), Hemeoxygenase-1(HO1) in addition to FTH1 in Hepatocellular Carcinoma (HCC) [62]. Subsequenly validation has been done in plethora of studies with regards to genes correlated with ferroptosis for instance Ferroportin (FPN), GPX4, Small Heterodimer Partner (SHP) along with Peroxisome Proliferator Activated Receptorγ (PPARγ) [62,95]. Taken together Nrf2 has been observed to control ferroptosis via three pathways i) iron metabolism [96], ii) lipid metabolism [97], along with ii) intermediatory metabolism [98]. Thereby Nrf2 has been isolated in the form of a significant controller as well as favorable for targeting of DKD. Fujiki et al. [99], observed that blockade of activin receptor like kinase 4/5(ALK4/5, that represents a TGF-β1 receptor) is capable of activating Nrf2 signaling to hamper erastin stimulated ferroptosis in HK2 cells [100]. In case of NRK-52E cells, TGF-β1 stimulation substantially resulted in upregulation of p53 as well as nuclear Nrf2 protein quantities, that was commensurate with the tendency of erastin stimulated cells in addition to animal models, pointing that Nrf2 is associated with ferroptosis in case of diabetic situations [57]. NRF2/FPN1 pathway activation is capable of diminishing myocardial - Ischaemia Reperfusion Injury (IRI) in diabetic rats [100]. Hampering of Nrf2 escalated the sensitivity of RTEC’s to ferroptosis resulting from escalated glucose, along with Nrf2 upregulation reverted the ferroptosis associated protein for attenuating cell damage in case of diabetic situations as per Li et al. [63]. HMGB1 possesses the capability of controlling ferroptosis via the Nrf2 pathway in mesangial cells in reaction to escalated glucose [60]. Thus Nrf2 action is incapable of being separated from ferroptosis. homeostasis. ferroptosis expression of Nrf2.
HO1: HO1 is implicated in catalysis of rate restricting step catabolism with regards to haem transformation to biliverdin, Carbon monoxide (CO) as well as iron. A plethora of studies have illustrated that HO1 / biliverdin /CO is implicated in the propagation of anti-inflammatory in addition to antioxidant stress [101]. In case of DKD, escalated quantities of HO1 stimulated by Nrf2 or Hypoxia Inducible Factor 1α (HIF 1α), efficaciously diminished ROS production along with ameliorated kidney damage [101,102]. Upregulation of expression of HO1 have been observed in studies in RTEC cells by ferroptosis stimulators [103]. Nevertheless, it continues to be uncharted if escalated HO1 stimulated in the form of reaction conferring protection or aggravates ferroptosis. Feng et al. [104], posited that diabetic renal tubular damage as well as fibrosis in case of db/db mice resulted in escalated quantities of HIF 1α in addition to HO1, that resulted in iron accrual in renal proximal tubule by downregulating ferritin. HO1 hampering ameliorated iron accrual, therefore avoidance of lipid peroxidation diminished ROS development [103]. Nonetheless Adedoyin et al. [103], observed that HO1 resulted in repression of erastin stimulated ferroptosis of RTEC cells in their study [103]. Intriguingly in case of Contrast Induced Nephropathy (CIN) model, diabetic rats had delivery of intravenous iopromide, haem hampered ferroptosis by activation of HO1/ Nrf2 along with upregulation of GPX4. Gao et al. [105], posited that HO1 as well as Nrf2 pathway comprised of a mutually positive buttressing cycle leading to self-underpinning in addition to sustenance of antioxidant capability [105]. It is debatable till now, is part of HO1 might be based on exhaustive controlling of iron quantities along with antioxidant actions What continues to be a hurdle is HO1 generating cytoprotective actions against kidney damage in case of DKD. Thereby, greater scientific research is imperative to estimate d the precise working of HO1 in DKD.
HMGB1: HMGB1 represents a nuclear protein, having universal expression, that is implicated in the propagation of stabilization of nucleosome in addition to DNA binding. It has been observed to work in the form of proinflammatory cytokine in plethora of pathologies for instance, cancer sepsis along with IRI [106]. Breaking the communication of HMGB1 as well as its receptor was illustrated to result in avoidance of circumstance in addition to formation of DKD [107]. Furthermore, HMGB1 was portrayed in the form of innovative modulator of ferroptosis by controlling Rat Sarcoma (RAS)/ c- Jun NH2-terminal Kinase (JNK) /p-38 signaling pathways in addition to a plausible target for leukaemia treatment [108]. A recent study illustrated that repressing HMGB1 possesses the capability of ameliorating escalated glucose stimulated ferroptosis in glomerular mesangial cells along with mitigates the propagation of inflammation. Wu et al. [60], further observed that HMGB1 Knock Out (KO) hampered the escalated glucose stimulated Toll Like Receptor 4 (TLR4)/ NFκB signaling pathway as well as the expression of Nrf2 in addition to its downstream targets for instance HO1, NQO1, Glutamate- Cysteine Ligase Catalytic subunit (GCLC) in addition to modifier of Glutamate- Cysteine Ligase (GCLM) [60]. Collectively, these observations suggest that HMGB1 is of considerable significance in ferroptosis controlling at the time of start along with propagation of DKD as well as apparently has been an ignored topic of crosstalk amongst ferroptosis in addition to inflammation in DKD.
ZRT/IRT like protein 14 (ZIP14): ZIP14 represents a divalent iron import protein that has organization in the proximal along with distal tubules, which pick up non transferrin bound iron from lumen [109]. Iron accrual is capable of escalating lipid getting deposited in the intestinal epithelial cells. Noticeably, upregulation of ZIP14 as well as FPN possess a significant part in the event [110]. Escalated quantities of ZIP14 have been in the proximal in addition to distal tubules, in case of focal segmental glomerulosclerosis along with IgA nephropathy that get aggravated by cell oxidative injury [141]. Hepatic ZIP14 hampering in case of TFR particular KO attenuated ferroptosis modulated liver fibrosis [111]. Wu et al. [112], found that ZIP14 might be implicated in the lipid getting deposited as well as stimulate ferroptosis in DKD rats in addition to hampering ZIP14 expression in HK2 cells possesses the capability of repressing escalated glucose stimulated ferroptosis [112].
N-acetylcysteine (NAC): N-Acetylcysteine (NAC), portrays a sulf hydryl group agent that is present in acetylated kind of L-cysteine. NAC portrays a precursor of the reduced GSH, that illustrates direct or indirect antioxidant characteristics [113]. Recent observations illustrated that NAC conferred protection against diabetic kidney damage by hampering OS [113]. A study with regards to myocardial - Iischaemia Reperfusion Injury (IRI) in case of diabetic rats, N-Acetylcysteine (NAC), ameliorated ferroptotic cell demise in addition to oxidative damage [114]. In case of a recent study performed in a diabetic beatle dog (stimulated by STZ & alloxan), combination of NAC along with insulin mitigated kidney tissue injury by repression of ferroptosis. Subsequently greater evaluating corroborated that NAC basically escalated the expression of GPX4 by upregulation of SIRT3 as well as hampering acetylation of Superoxide Dismutase (SOD), thus hampering cell ferroptosis along with improvement of working of DKD kidney [115].
AMP-activated protein kinase (AMPK): AMPK in the form of central controller of energy homeostasis has been broadly explored in plethora of human diseases, as well as has gathered escalating attraction in the form of an intervention targeting DKD. Downregulation of expression of AMPK was found in renal tissue in High Fat Diet (HFD) stimulated obesity rats [117]. In case of energy stress, for instance glucose starvation in addition to ATP Lee et al. [44], observed that activated AMPK is critical for hampering erastin stimulated ferroptotic cell demise [44]. It got corroborated that AMPK hampered PUFA generation by hampering AcetylCoA Carboxylase (ACC) actions. AMPK activated ACC implicated in catalysis of generation of malonylCoA from acetylCoA, the rate restricting enzyme in FA generation. Akin to that AMPK has been determined in cardiomyocytes via separate pathway. In this study AMPK was corroborated to hamper ferroptosis stimulated by IRI in diabetics through inactivating NADPH oxidase (NOX2), pointing that hampering actions of AMPK/ NOX2 on ferroptosis might be one more protection conferring mechanistic modes for Myocardial Infarction (MI) hearts [114]. In db/db mice earlier studies observed that AMPK activation possesses the capability of diminishing ROS generation along with escalating mitochondrial bio generation as well as ETC actions to ameliorate kidney injury in diabetic models [116]. Till now direct validation with regards to actions of AMPK on renal cell ferroptosis in case of diabetic situations is inadequate/ lacking. Nonetheless, it merits assessment if AMPK possesses positive influence on DKD by hampering ferroptosis.
Other controlling factors: Apart from controlling mechanistic modes variable other mechanistic modes have been recently displayed. For instance, Jin et al. [117], observed that murine Vezf 1 gene, mmu_circ RNA_0000309 is capable of upregulating GPX4 by competitively binding miR 188-3p, that ameliorated mitochondrial in addition to podocytes injury by hampering ferroptosis. Nevertheless, hsa_circ RNA_0000309 is separate from mmu_circ RNA_0000309. In reference to clinical applications human DKD there is requirement for the identification of candidate genes [117]. In case of type 1 diabetic mice as well as escalated glucose cultured HK2 cells, it got illustrated that Circ _ASAP2 by binding miR 770-5p diminished inflammation in addition to ferroptosis by SRY related high mobility group box transcription factor (SOX2) /SLC7A11 pathway [118]. Peroxiredoxin6 (PRDX6) a newly isolated member of the antioxidant enzymes family, works in the form of a negative modulator of ferroptosis [119]. Certain studies further corroborated PRDX6 was capable of modulating Lipopolysaccharides (LPS) stimulated AKI, by diminishing OS along with inflammation [120]. Zhang et al. [58], observed that Prdx6 expression got controlled by Specificity Protein (Sp1) at the level of transcription, as well as upregulating Prdx6 possesses the capability of avoidance of, podocytes dysfunction in DKD via mitigation of OS in addition to ferroptosis [58]. Salusinbeta, portrays a bioactive peptide comprised of 20 amino acids whose translation takes place from alternatively splicing of mRNAs of the torsion dystonia related gene (TOR2DA) [121]. Salusinβ takes part in escalated glucose stimulated ferroptosis based on Nrf2 in HK2 cells [122]. In case of Bioinformatics analysis, it was demonstrated that ferroptosis was implicated in significant enrichment of biological events in tubulointerstitium [123]. Screening hub genes was performed by Hu et al. [159], dependent on Weighted Correlation Networks Analysis (WGCNA) algorithm as well as isolated intercommunicated transcription factors in addition to non-coding RNAs (nc RNAs) utilization of principal component analysis. They observed that FPR3, CD3AR1, CD14, ITGB2, RAC2, ITGAM were the hub genes associated with DKD initiation [124]. Correspondingly ALOX15 manipulated lipid was determined in intrinsic glomerularcells through Bioinformatics analysis. miR 142 along with miR650 might be implicated in controlling ALOX15 [69]. Urinary metabolomics in a recent study displayed metabolic changes in DKD, inclusive of ferroptosis of signaling pathway [125]. Escalating evaluations have isolated genes correlated with utilization of Bioinformatics methodologies in addition to guiding for the asssessment of the particular mechanistic modes by further analysis.
Pharmacologic studies associated with ferroptosis in DKD
Recognized the manner ferroptosis possesses part in the pathological events of DKD, ferroptosis is an upcoming target for the avoidance as well as propagation of kidney injury. Here we detail the propagation in reference to experimental agents targeting ferroptosis in diabetic animal / cell models, which corroborate that repressing ferroptosis possesses remarkable plausibility for the DKD treatment.
Antihyperglycemic agents
Antihyperglycemic agents are advantageous for the working of the kidney by diminishing albuminuria in addition to postponing propagation of kidney fibrosis in DKD [3]. Nevertheless, greater studies have observed that they have the capacity of ameliorating kidney injury by hampering ferroptosis.
Thiazolidenediones (TZD) have the capacity of diminishing blood glucose quantities along with basically escalate the insulin sensitivity [126]. Additionally, in the form of ACSL4 hampering agents, TZD are capable of decreasing the plasma free FA quantities. It was displayed that rosiglitazone was the maximum robust ACSL4 hampering agent [127]. Wang et al. [56], corroborated that rosiglitazone was capable of diminishing blood glucose as well as urinary albumin by hampering ferroptosis in addition to inflammation in case of both type 1 diabetic along with type 2 diabetic mice [56]. Fenofibrate is implicated in the controlling of lipid as well as activating PPAR’s. Apart from diminishing blood lipid quantities, it is capable of causing improvement of renal injuries in addition to proteinuria resulting in view of hyperglycemia [128]. Fenofibrate was confirmed to influence ferroptosis associated signaling pathways for protection conferred to the working of the kidney in DKD, along with Nrf2 was corroborated to be of considerable significance in the event of amelioration [63]. SGLT2 hampering agents diminished glucose reabsorption in the proximal tubules via SGLT2 to facilitate hypoglycaemic actions [129]. Quagliariello et al. [130], displayed that Empagliflozin (EMPA) possesses the capability of causing improvement of cardiac working by diminishing ferroptosis via NLRP3 inflammasome as well as myeloid differentiation innate immune adaptor gene (MyD88) associated signaling pathways in mice that got treatment with Doxorubicin (DOX) [130]. Additionally, it got Demonstrated that Dapagliflozin (DAPA) was capable of improvement of renal working in DKD by repression of ferroptosis by SLC40A1 stabilization. The mechanistic modes might be DAPA binding FPN resulted in hampering of ubiquitination breakdown of FPN [131], yielding an innovative therapeutic manner of clinical applications DAPA. Liraglutide portrays an inherent human Glucagon Like Peptide1 Receptor Agonists (GLP-1RA) [132], is capable of activating the Nrf2/ HO1 signaling pathways to ameliorate Hepatic fibrosis in db/db mice by provision relief from ferroptosis [133]. Apart from that, it possesses the capability of attenuating dysfunctional cognition of diabetic mice by decreasing ROS as well as iron getting deposited [134]. Nonetheless, part of Liraglutide continue to be uncharted.
Taken together it is not tough to observe that the actions of antihyperglycemic agents on hypoglycaemia or protection conferred to the kidney is intricately associated with iron metabolism, Nrf2 in addition to lipid metabolism as well as canonical mechanistic modes. There by, it portrays that ferroptosis is of considerable significance in the propagation of DKD.
Agents causing hypotension: In view of diminished Glomerular Filtration Rate (GFR), metabolic conditions in addition to activation of renin-angiotensin-–aldosterone – system (RAAS hypertension usually associated with DKD. A plethora of agents causing hypotension have the capacity of decreasing the urinary albuminuria along with halt the propagation of kidney diseases [4]. Nevertheless, germanely occasional studies exist on, if ferroptosis portrays a therapeutic target of agents causing hypotension in DKD therapy.
Entresto alias LC696 gets comprised of sacubitril as well as valsartan that got recommendations, Food and Drug Administration (FDA) Heart Failure (HF) therapy in the clinical scenario [135]. An escalating quantity of Clinical trials confirmed that Entresto is attractive for protection conferred to the kidney as well as decreasing hypertension. Recently Zhao et al. [94], displayed Entresto was capable of hampering ferroptosis to diminish DOX stimulated cardiotoxicity. In the meantime, it was observed that such observations displayed that it had the capacity of repression of ferroptosis activating by (SIRT3 pathway for decreasing the injury to myocardium, yielding an innovative therapeutic manner of clinical applications of Entresto in DKD [136]. Utilization of captopril, a canonical antihypertensive agent from the family of ACE hampering agents has got done for decreasing the urinary albuminuria for over last decade [137]. As per current availability of studies captopril was incapable of controlling ferroptosis. In case of murine model study of acute radiation syndrome, iron getting deposited in the spleen was not attenuated, in addition expression quantities of SLC7A11 continue to be unaltered subsequent to captopril therapy [138]. Additionally, Ca2+ channel blockers have the capacity of modulating iron overload through divalent metal transporter 1 [139], nonetheless the availability of Ca2+ channel blockers in repression of ferroptosis continue to be unaltered. Thereby, greater corroboration is required in depth evaluation of part of antihypertensive agent in controlling ferroptosis.
Canonical herbal agents: Recent studies have found that a plethora of herbal constituents possess advantageous action in repression of ferroptotic cell demise in DKD. Germacrone, a bioactive component of Rhizoma Curcuma was observed to result in improvement of ferroptosis stimulated kidney injury in db/db mice through the ROS / GSH/ GPX4 axis. In case of MPC cultured in escalated glucose, hampering ferroptosis attenuates escalated glucose stimulated podocytes mitochondrial injury along with mmu_ mmu_circ RNA_0000309 possesses considerable significance part in such an event [117]. Quercetin, which portrays a flavonoid possesses broader organization in nature, has been confirmed to diminish the risk of T2D as per epidemiological asssessments [140], as well as it does so by plethora of mechanistic modes, that includes antioxidant, anti-inflammatory, anti-fibrotic actions [140]. Li et al. [141], observed that quercetin possesses the capability of repressing ferroptosis of pancreatic β cells in generation of T2D [181]. Concomitantly, Jiang et al. [142], observed that quercetin was capable of decreasing the hepatic lipotoxicity in case of HFD stimulated mice by downregulation of mitochondrial ROS modulated ferroptosis [142]. Additionally, quercetin has the capability of protective effects in AKI by decreasing the expression of Activating Transcription Factor (ATF3) [143]. Nevertheless, greater corroboration is required with regards to quercetin working in DKD stimulated ferroptosis. Salidroside portrays a constituent of the canonical Chinese medicine obtained from Rhodiola RoSeal, that has been displayed to recontour Gut Microbiota (GM) [144], in addition to restrict the accrual of iron [145]. In case of db/db mice, Shi et al. [146], observed that salidroside possesses the capability of diminishing blood glucose quantities, whose controlling might result from altering GM along with controlling of escalated glucose iron metabolism [146]. Calycosin (C16H12O5) portrays an isoflavone root stem extract of Astragali Radix, that possesses anti-inflammatory, immunocontrolling, antioxidant as well as anti-viral characteristics [146]. In case of db/db mice, in addition to HK2 cells cultured in escalated glucose, calycosin hampered escalated glucose stimulated ferroptosis by influencing GPX4 along with NCOA4 quantities [147]. Glabridin portrays, a polyphenolic flavonoid along with is the bioactive constituent of licorice, that has been demonstrated to possess hypoglycemic as well as protective role in T2D [148]. Tan et al. [149], observed that glabridin was capable of resulting in restoration of antioxidant system injured due to escalated glucose, by diminishing iron quantities in DKD in addition to recovery of quantities of ferroptosis correlated biomarkers. They posited that such anti-ferroptosis actions of glabridin were modulated by the Vascular Endothelial Growth Factors (VEGF) /Akt/ Extracellular signal –Regulated Kinase (ERK1/2 pathways [150]. Platycodin D root extracts from Platycodon grandiflorum represents a triterpenoid saponin that possesses plethora of pharmacologic characteristics inclusive of anti-tumor anti-viral, anti-inflammatory, antigeing along with neuroprotective actions. It got displayed that Platycodin D possesses the capability of hampering escalated glucose stimulated ferroptosis in case of HK2 cells by resulting in GPX4 upregulation [151]. Schisandrin A that represents lignans, identified from Schisandra chinensis SCE fruits [152, rev by us in ref 153) have been shown to be used for longstanding time in canonical Chinese medicine. It has the capability of hampering ferroptosis as well as conferred protection against kidney injury by diminishing ROS ameliorating inflammatory reactions as well as attenuating mitochondrial injury via adiponectin receptor 1 (ADIPOR1) / AMPK / Nrf2 /HO1/ GPX4 pathways in HFD/ STZ stimulated diabetic mice [152]. 7 hydroxycoumarin (alias umbelliferone), represents a coumarin derivative that possesses broader organization in umbelliferae plants, that has plethora of actions for instance anti-inflammatory, antihyperglycemic actions. It has the capability of hampering ferroptosis by activating Nrf2 /HO1 pathways in case of db/db mice in addition to HK2 cells stimulated by escalated glucose, thus avoids propagation of DKD [154]. Figure 4 summarizes in a nutshell the full iron metabolism with role of System x, stimulators & inhibitors of ferroptosis [rev in 155]. Li T, & Yu C [152], further described part of cuproptosis in renal fibrosis.
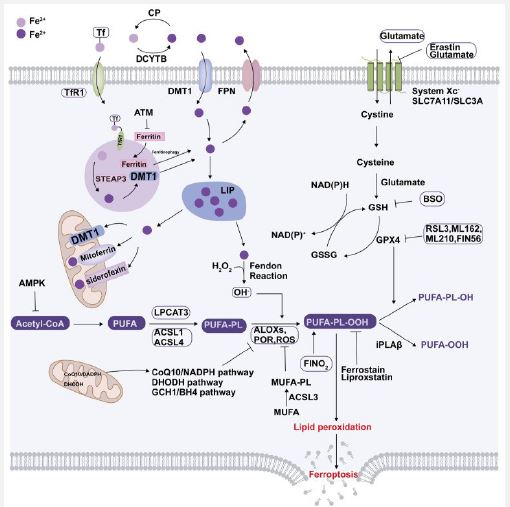
Thus here we have comprehensively provided a narrative review with regards to role of ferroptosis in case of DKD inclusive of iron overload, diminished antioxidant capability as well as accrual of lipid peroxides. Furthermore, we stressed on the way ferroptosisis is associated with DKD with the advancements made in this field with highlighting ferroptotic modulators of DKD. Subsequently we have dealt with the pharmacologic advancements targeting ferroptosis in case of DKD with the idea of generation of newly isolated approaches for the DKD treatment.
Escalated proof has illustrated that ferroptosis possesses critical in the initiation in addition to propagation of DKD along with hampering ferroptosis protection conferred to the working of the kidney in DKD [56,63]. Nevertheless, certain noticeable botherations have to be taken into account i)The precise mechanistic modes that stimulate ferroptosis continue to be uncharted. Despite practically all scientific researchers have agreement on accrual of iron in the RTEC [52], whether iron overload, is the one that is implicated in guiding accrual of ROS by numerous pathways? Additionally, the downstream factors of lipid peroxidation continue to be uncharted. ii)Greater asssessment to display ferroptosis in DKD, particularly in case of coexistence of myriads of pathological situations accompanied by DKD as well as in case of variation of factors mutually influence everyone. Apart from that, DKD occurs by modulating plethora of cell demise pathways for instance autophagy [67]. Nonetheless, the manner such cell demise modalities control ferroptosis in the generation of DKD has requirement of greater evaluation. iii) The present ferroptosis correlated biomarkers for the anticipation of ferroptosis in pre-clinical studies are no particular for instance GPX4 in addition to ferritin [25,37]. The generation of proteomics along with metabolomics methodologies has escalated finding of plausible biomarkers with regards to the pathogenesis as well as role of ferroptosis in DKD [125], however, following experiments in diabetic animals in addition to clinical validation is required before human applications merits greater time [125]. Furthermore, the manner elaborated earlier antihyperglycemic agents have the capacity of hampering ferroptosis, yielding an innovative therapeutic manner of clinical applications in for instance AKI [156]. Despite, selective hampering agents of ferroptosis have confirmed extensive improvement in diabetic animals / cell models, though human clinical trials have yet to be conducted using ferroptosis particular agents that need urgent asssessment. Greater work has been ongoing, recently Xia et al. [157], displayed the role of the transcription factor CCAAT/Enhancer Binding Protein Alpha (C/EBPα) in DKD pathogenesis. They found that C/EBPα upregulated ACSL4 expression by binding to its Transcription Regulatory Sequence (TRS), leading to escalated elevated lipid peroxidation as well as ferroptosis. Additionally, hampering or genetic ablation of C/EBPα ameliorated ferroptosis in addition to abrogated tubular damage in DKD. These outcomes emphasized the C/EBPα-ACSL4-ferroptosis pathway in the form of an attractive therapeutic target for DKD treatment. Additionally, Zhang et al. [158], assessed the actions of atorvastatin on diabetes stimulated renal injury along with ferroptosis signaling in a mouse model of diabetes stimulated by STZ as well as HFD with/ without atorvastatin. Atorvastatin significantly diminished plasma lipid profile, proteinuria, renal injury in diabetes mice. Atorvastatin diminished renal ROS, iron accrual in addition to renal expression of Malondialdehyde (MDA), 4-Hydroxynonenal (4-HNE), Transferrin Receptor 1 (TFR1), along with escalated renal expression of GPX4, NRF2 as well as Ferritin Heavy chain (FTH) in diabetes mice. Commensurate with the observations in vivo, atorvastatin resulted in avoidance of escalated glucose- stimulated ROS generation along with Fe2+ accrual, an escalated expression of 4-HNE, MDA as well as TFR1, in addition to a diminished cell viability along with the expression of NRF2, GPX4 as well as FTH in HK2 cells. Atorvastatin further reverted ferroptosis inducer erastin- stimulated ROS formation, intracellular Fe2+ in addition to the alterations in the expression of aforementioned ferroptosis signaling molecules in HK2 cells. Apart from that, atorvastatin ameliorated escalated glucose- or erastin stimulated - mitochondria damage. Ferroptosis hamperor, ferrostatin-1 along with antioxidant N-Acetylcysteine (NAC) equally reverted the expression of escalated glucose- stimulated ferroptosis signaling molecules. Their data embraced the belief that statins possess the capability of hampering diabetes stimulated - renal oxidative stress as well as ferroptosis, which might aid in statins protection of diabetic nephropathy [158].
